
放射線の低減に土壌微生物が関係する新たな知見
- このフォーラムに新しいトピックを立てることはできません
- このフォーラムではゲスト投稿が禁止されています
投稿ツリー
-
 放射線の低減に土壌微生物が関係する新たな知見
(go, 2013/9/23 4:30)
放射線の低減に土壌微生物が関係する新たな知見
(go, 2013/9/23 4:30)
-
データ分析表の説明
(go, 2013/9/25 9:14)
-
光合成細菌はなぜ放射線を吸収できるのか?
(go, 2013/12/31 8:49)

go
 投稿数: 125
投稿数: 125
投稿数: 125
●要点
これは、「土壌微生物の働きによって放射線量がどのように低減するか」という実験報告である。
・実験期間は2013/5/6~9/19、放射能の漏入漏出のないミニハウス内のプランターにて行い、
プランター土壌表面およびプランター底面の放射線量を測定した。
・データ処理および分析は、連続測定20個を平均し、空間線量で補正し、
微生物の働きによる放射線量の変化を浮き彫りにるすため、放射能半減期崩壊による減少分を補正し、
さらに直近7日間を移動平均処理することによって放射線特有のデータバラつきを平準化した。
その結果は次のとおり。
①土壌から発するγ放射線は、微生物の働きで低減することが明らかになった
②γ放射線の低減は光合成細菌の働きによるものと思われる
③放射線の低下の割合は、底面よりも土壌表面の方が大きい
④ひとたび微生物が働いた後には、土壌が乾燥しても土壌表面の放射線低減力は残っている
⑤光合成細菌を含む微生物を使って人為的に放射線を軽減することができる
⑥微生物の働きによる「放射能崩壊加速説」の信ぴょう性が高まった。
-----------------------------------------------------------------------------------------
既に「放射能低減実験その2」にてライブ的に経過を報告してきたが、多少の誇張・推測・勘違いもあり、
改めてこの場で実験全体を整理し考察し直した。
なお、実験が未熟だった放射能低減実験その1 も参考まで。
-----------------------------------------------------------------------------------------
1.実験の目的とその背景
(1) 自分の手で土壌微生物が放射能を低減する客観的なデータをつかみ、
その事実詳細を誰にでも分かりやすい形で公表すること
・放射能汚染や風評で苦しんでいる東北・北関東および健康不安を抱いている全国の生活者のみなさんに、
・誰にでも簡単にできる「放射能汚染対策の可能性」を知って欲しく、、
・家族を守るためにも、自宅や地域等にて放射能対策の活動をしていきたいものと願っている。
(2) 家族の健康や食の安全を国や行政だけに任せてはいけない
・チェリノブイリ原発事故のあと、農地にEM(有用微生物群)を施すことによって、
放射能が低減できることが実証実験で報告されていた。
・植物への放射能吸収が抑えられたこと
・農地の放射能が低減したこと
・また、フクシマの高度に放射能汚染された地区で、EMなどで有用発酵した肥料を使った農業では、
・植物に吸収される放射能はND(検出限界外)であったこと
・EMを散布した短期間に土壌の放射能が低減したこと
など、幾多も報告されている。
・しかし、国や行政や多くの御用学者やメディアまでもが、このような微生物による放射能低減効果の
事実を認めようとしないばかりか、微生物を拒絶または忌み嫌う姿勢が続いている。
放射能汚染で苦しむのは、汚染地域居住者や子どもや健康に不安を持つな生活者であり、
農林漁業従事者をはじめ、風評被害の産業である。
国は、除染に莫大なお金を投入しながら、環境回復のための基本的な放射能対策の指導ができないまま、
住民にも国民にも世界に対しても、深刻な事態を軽視し、安全だと言い続けて責任を逃れようとしている。
この日本を、どうしようもない国にしてはならない、生活者が安心して暮らせる国にしていきたものである。
2.実験の準備
放射能低減実験は、平成25年5月連休で実験環境を用意し、5月8日から6月16日まで「実験その1」を行い、
6月26日から実験方法を新たにして今回の「実験その2」が始まった。






放射能汚染土に混ぜた微生物関連資材や実験経緯などは、別トピック「放射能低減実験その2」 に
ライブ的にUPしてきたのでご覧ください。
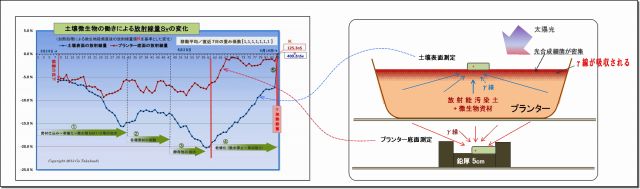
3.計測方法とデータ表
(1) 実験の概要
(実験方法) 放射能汚染土に微生物資材を混ぜたプランターにおいて、
土壌表面およびプランター底面から放射される放射線量を計ること
(実験者) 高橋剛(このサイト運営者)
(実験場所) 岩手県紫波郡矢巾町赤林(農村地帯)に設置したミニハウス
(実験期間) 平成25年6月26日~同9月19日(85日間)
(微生物叢環境づくり資材) モミ殻燻炭、ピートモス、培養土、EMボカシ、
EMセラミックパウダー、EM活性液、EM3(光合成細菌強化EM)など
(植えた植物) ヒメイワダレ草
(実験データ) プランター底面とプランター土壌表面の2ヶ所の放射線量を計り、
各々連続する20データを平均した
(求める分析データ) 微生物群などの投入や環境変化に応じた放射線量の変化
(2) Excelによるデータ表は次のとおり
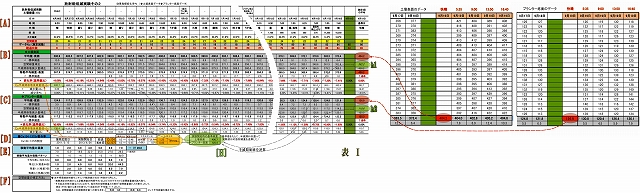
(表の構成)
・左側は、総括表である (中間時期を割愛し、実験初期と実験終期と並べた)
・右側は、別シートに収めた計測データ表の終期だけのもので、
最終日は時間をずらした計測値4回分の平均を求め、それを総括表にリンクしている。
詳しくは、子スレッドにて 「データ分析表の説明」 としてUPしている。
4.実験の経過
(1) 実験は85日間、土壌表面およびプランター底面から発生する放射線量を測定
・データ測定は、ほぼ毎日1回測定し、連続20個のデータを平均化し、それを元に、
・外から飛び込んでく空間線量を補正し、
・さらに汚染土に含まれる放射能自体の物理的半減期崩壊による減少値分を補正することで、
微生物だけの影響による放射線量の変化を知る というものである。
・そして、バラつきある直近7日間のデータを移動平均処理にて平準化した。
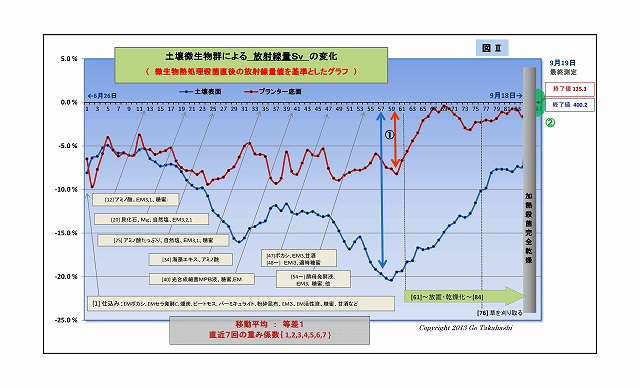
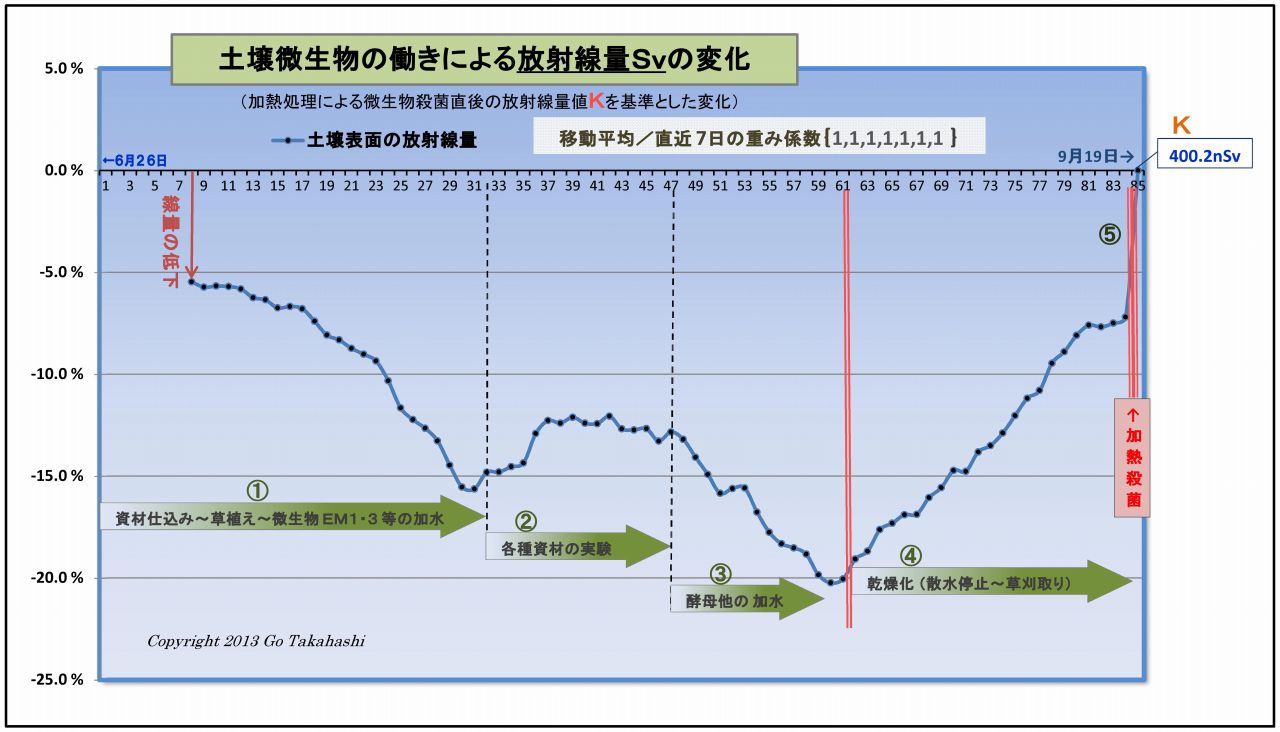
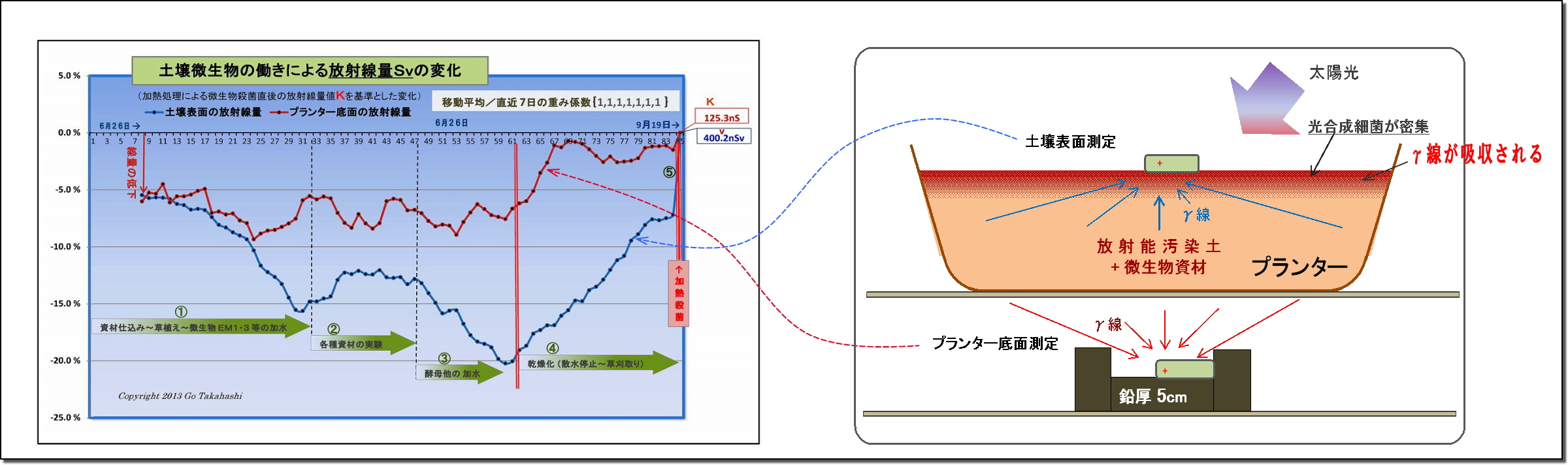
・下図は、このようにして求めた放射線量Svの変化をグラフ化したものである
・変化の基準としたデータは、微生物が含まれない汚染土の放射線量値である。
(実験最終段階で実験土壌を加熱殺菌して測定した)
・グラフの大まかな見方は、
・ヨコ軸は85日間の時間軸、タテ軸は基準値からの変化%である
・図中の①に示す幅の大きさが「微生物群の働きによる放射線の減少量」 である。
・スタート時点で、すでに-6%以上も下がっている理由は、
土壌には「実験その2」以前から含まれている微生物が、すでに働いていること、
そして、この実験開始時に、更に新たな微生物関連資材を加えためである。
・また、グラフは直近7日間の移動平均した値のものなので、放射線独特のバラつきは平準化されているが、
実験直後は直近データが足りないために、初日から数日間は平準化しきれず生データに近いものとなっている。
・なお、グラフの右後半61日目から放射線量が上昇し続けた理由は、
実験終盤の目的の一つである「意図的に土壌の乾燥を促した」したもので、
実験当初の放射線量に比べて増えている理由は、「放射線の減少量(図中の①)が減った」 ということである、
決して「微生物によって放射線量は増えたということではない」 ことを強調しておく。
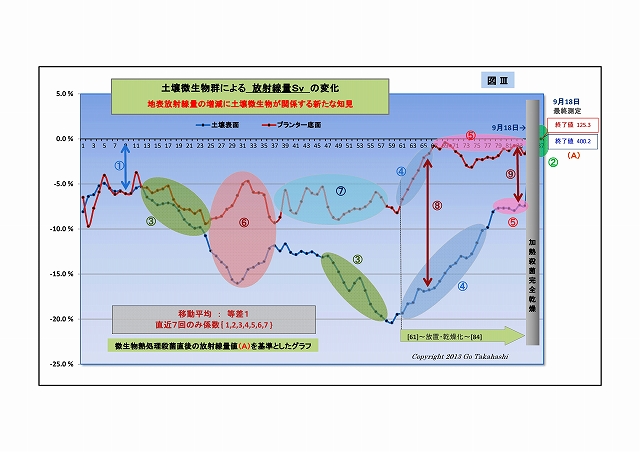
(2) 実験は、大きく3フェーズになっている
[第一フェーズ] 初日~31日目
・開始から2週間目頃から、放射線量が少しずつ下がってきた。
・確実に下げたいがために、微生物叢を豊かにするためのいろんな資材を適時に散布した。
特に光合成細菌が増えて活性化できる環境づくりを強く意識した
・しかし、25日目から、底面放射線が上昇に反転し、
続く31日目からは、表面放射線量も低下が止まり反転上昇した。
・それまで順調に低下したものが、なぜ反転上昇したのか、分からずに頭を抱えてしまった。
[第二フェーズ] 32日目~60日目
・しばらくは上昇が続いたが、光合成細菌にとって有用と思われる様々な資材を試行錯誤で
散布はしてみたが、放射線量をコントロールしかねていた。
・しかし、これまでは光合成細菌だけに目を向けていたけれど、酵母菌や乳酸菌などの共生菌を
整えてみよう思い、47日目に自家製の甘酒の希釈液など酵母菌を増やす資材を散布したら、
表面および底面の放射線量が再び勢いよく下降しはじめた。
・この頃になると、ヒメイワダレ草は勢いを増してきている。


[第三フェーズ] 61日目~最終85日目
・60日目になると、またもや土壌表面放射線の下降が止まり上昇し始めた。
・これをキッカケに、実験終盤のテーマとしていた「土壌の乾燥(微生物の活性を落す)」 に進んだ結果、
・予想通り、放射線低減量が減りつづけ、平衡に達しようとしていた。
乾燥をもっと加速させるべく草を刈り取ってはみたが、切った茎から新芽が出てくる始末、
プランターであっても環境空気中に湿度がある限り完全に乾燥するのは無理だと悟った。
(なお、これ以降、刈り取った草も含めて測定している)
そして最後の手段、微生物を加熱殺菌処理して、汚染土そのものの放射線量を測定した。
・微生物の働きを完全の停止するには、加熱殺菌が確実であろうと判断し、
台所のハイカロリーコンロで中華鍋で土を焼いて微生物を加熱殺菌し、
(少なくとも微生物の細胞タンパクは破壊し光合成機能は喪失したと思う)
・それをプランターに戻して翌日に最後の測定を行った。
・時間をあけながら4回測定し、
・その平均値を最終日の測定値とする
この値が「微生物に影響を受けない汚染土の放射線量」なので、この値を分析やグラフ化の基準値とした。









5.考察
まず最初に、この実験は、放射線測定器があれば、誰でも容易に試すことができることを強調しておく。
また、分析データは、次のよう処理によって、公開に耐えるに足りる信頼性があるものと思っている。
・1回の測定データ連続23個のうち、最初の3個を捨て、残り20個を平均していること
・平均値に含まれる空間線量(天候によって変動)を補正していること
・更に、日時の違いによってバラつき気味な値を、移動平均処理にて平準化していること
(1) 土壌から発するγ放射線は、微生物の働きで低減することが明らかになった
・グラフ全体から、明らかに微生物の働きによって放射線量が変化することが分かった。
・土壌微生物叢を、外部から人的にコントロールすることで、土壌放射線量を低減できることが証明された。
・従来の放射能に対する科学者たちの常識では、
・γ放射線の透過力はとても強く、金属をも通過するので、
・微生物ごときで放射線は弱まるハズがない!
と、いうことであった思われるが、
これが、いとも簡単に否定されることが実証された(新知見)
(2) γ放射線の低減は光合成細菌の働きによるものと思われる (微生物による放射線吸収説)
・光合成細菌は、光(電磁波)エネルギーを吸収して光合成を行う微生物である。
γ線も電磁波の一種であり、放射線を当てることで光合成細菌は増殖することも分かっている。
・放射能γ線エネルギーは、光合成細菌の細胞内の光合成メカニズムによって利用・吸収されるので、
吸収された分だけγ線エネルギーは減衰し、光合成細菌の密度が高いほど減衰量は大きくなる。
放射線の減衰とは、つまりは 「放射線の低減」 のことである。
(3) 放射線の低下の割合は、⑧に見られるように、底面よりも土壌表面の方が大きい
・これは、表面側の方が減衰量は大きいことを意味している
光合成細菌は、土壌の深場より表面近くの方が増殖しやすく密度が濃くなっている、と思われる。
・表面と底面の格差は、微生物叢が豊かになっているのだろか、時間経過とともに大きくなっている。
・しかし、④⑤に見られるように、土壌の乾燥が進んでも明確な差は残っている。
(4) 一たび微生物が働いた後には、土壌が乾燥しても土壌表面の放射線低減力は残っている
・乾燥が進んだ④~⑤を見ると、底面び方は微生物の働きがほぼ止まったのに対し、
表面の方は、低減が-7%ほどで止まったままになっているが、
・光合成細菌は土壌が極度の乾燥状態であっても、γ放射線を遮蔽し続ける能力が残っているものと思われる。
(5) 光合成細菌を含む微生物を使って人為的に放射線を軽減することができる
・フェーズ1の③に見られる放射線の低下は、光合成細菌をはじめとする微生物叢を豊かにするための
有効な資材を放射線汚染土に混ぜた結果であること。
・フェーズ2の③に見られる放射線の低下は、更なる低下を目指して、人為的に微生物叢をコントロール
した結果であること。
・この時、特に意図したことは、光合成細菌の働きを助けるために酵母菌や乳酸菌を補強したことである。
このように、人為的にγ放射線を減らすことができることが実証された。
(6) 微生物の働きによる「放射能崩壊加速説」が浮上した
・⑥に着目すると、「変化の方向が表面と底面で逆である」 この原因は何だろうか?
・それまでは表面・底面ともに放射線がムーズに低下したのは放射線吸収説で説明できる、
しかし25日目を境に突然底面だけが上昇方向に転じたが、
土壌表面は依然として下がり続けているので、光合成細菌は増え続けていると考えられるが、
だとすると底面方向も下がっていいものだが、実際の挙動は違っていた。
・考えられる一つは、
光合成細菌には鞭毛があって、光に向かって進む性質があるので土壌表面に移動したのではないか、
という見方である(光合成細菌移動説)。
しかし、⑥の後半などでは、その逆方向の動きも散見されるので、移動説だけの説明には無理がある。
・別の考えもある、
光合成細菌は「放射能崩壊を早める」という説がすでに囁かれていたものだが、
果たして、この現象がそうなのだろうか?
・そこで、移動平均の重み係数をいろいろと変えてグラフにした(下にUP)。
右端グラフ「重み係数なし(生データ)」をよく見ると、
・放射線量は激しく振れているが放射線特有のバラツキとは異なった挙動であること、
・表面と底面方動きの向きが反対になっている箇所が随所にあること、
・時には土固有の放射線(基準値)をオーバーする箇所も散見されること、
・そして⑦を見ると、表面の変動よりも底面の変動の方が激しいことなど、
放射能吸収説や光合成細菌移動説を併しても説明つかない挙動ある。
・ひょっとして、微生物叢の働きによる「放射能吸収」と「放射能崩壊加速=放射線増加」が、
並行して起こり、互いに拮抗している状況のように思えてならない。
・「放射能崩壊加速説」なら、半減期より早く崩壊する分だけけ放射能は低減される、ということだ。
・「放射能崩壊加速説」の裏付けには、実験前後で放射線Bq値が減少している証が必要があるが、
実験その2では、実験前に根拠となるデータを取っていなかったことが悔やまれる。
(すでに実験その3が進行しているので、その実験の終わりに、加熱殺菌して放射線量を計るつもりである)



6.雑感・提言
(1) 土壌の微生物叢を豊かにすることで、人体に悪影響をおよぼす放射線Svを減らすことできる
・自宅の庭や花壇や塀周りや地域の子どもたちのの遊び場など、緑を豊かにしましょう、
・地肌を出さないような管理をしたいものですね。
・放射線量は、10~20%は簡単に減らせると思います。
(2) 誰にでも放射線を低減できる方法
・すでに、福島をはじめ、宮城、栃木、茨城、群馬など各地の市民、ボランティ、農林畜産業の方々が、
自発的にEM(有用微生物群)を活用した射能対策を行って、成果をあげており、
EMの具体的な活用方法と成果事例が多くのサイトにUPされているので、それを参考にしていただきたい。
・しかし、必ずしもEMにこだわる必要はない。
光合成細菌はいろんなメーカから流通されているし、
米ヌカなどを散布して、その上から糖蜜などでしっかり乳酸発酵した培養液の希釈液を散布するだけで
良好な微生物叢が形成され、自然に光合成細菌が増えていくものである。
・私の実験でも、EMだけに頼ることはしていないが、
光合成細菌をはじめ、いかに土壌微生物叢を機能的に豊かにするかに手段は選ばなかった。
(簡単な放射能の低減方法の提案)
・やはり、EM発酵液は必須です、手軽に確実に乳酸菌・酵母菌が培養できるからです。
全国どこでも、お近くに培養して実践しているボランティアな人が必ずいますから、
一度訪ねてみることが早道です。
・そのEM希釈液に光合成細菌を少し足して、散布するだけです。
散布直前に米ヌカなどを薄く撒けば、ベターな微生物叢ができる。
・お庭などでは、土を丸出しにしないことです、
・放射能を含んだ粉塵が肺を直撃しますから、
・ぜひカバープランツを植えましょう、
芝生もいいけど刈込などの管理が面倒でしょうから、
ヒメイワダレ草がおススメです、
・繁殖力旺盛、根は深く張り、背丈は伸びず地面を這って広がる、踏まれても強く、雑草にも強い、
冬は表面は枯れるけど根はしっかりと生きていて、5月ころからは緑の絨毯にる、
・年中小さな花が咲いくれるのもうれしいし・・・。
(3) 研究者・科学者のみなさんにお願いしたい
・いまは、生活や環境活動や農業などに微生物を使った活動や実践は珍しいものではなくなった、
しかし、微生物による放射能低減に関する研究は非常に少ない現状にある。
私の実験のような結果は、誰にでも簡単に再現できるものであり、
再現性ある現象には、後日科学的に証明されてきたことことが科学の歴史でもあった。
・私の実験でも、放射能に及ぼす光合成細菌の働きにはナゾが多いことがわかった。
・放射線吸収し放射線を減衰させること
・放射能の崩壊を即しているのではないか?
・逆に、放射能の崩壊を抑えているのかも知れない?
・放射能核種の元素転換を起こしているのかも知れない?
・・・・・
ぜひ、研究して欲しい、科学的に証明して欲しい、その知見などをネット公開して欲しいと願っています。
(4) どこでも行われている 放射線量Bq測定方法は、果たしては正しい値を出せるのだろうか?
・放射能の強さ計るには、放射線を検知し、線量をカウントしてから、Bq,CPM,Svを導きだしている。
・ところが、私の実験で分かるように、土壌微生物の密度・活性度の違いや、
乾燥の度合、採取する深さの違いなどで、放射線カウント数も変わることが分かった。
・放射線Bqベクレル測定の方法を調べてみると、「土壌サンプルは乾燥すること」とは明記されていないのだ。
・土壌微生物の働きで放射線が弱まる、という認識が全くないいからだと思われてならない。
正しく計るには「土壌を加熱殺菌」すべきである、と考える。
--------------------------------------------------------------------------------------
ぜひ、私の実験方法と導いた結論めいたことなどを、一人でも多くの方々に追試していただき、
より普遍的な放射能対策の知恵として広がってもらいたいと念じています。
これは、「土壌微生物の働きによって放射線量がどのように低減するか」という実験報告である。
・実験期間は2013/5/6~9/19、放射能の漏入漏出のないミニハウス内のプランターにて行い、
プランター土壌表面およびプランター底面の放射線量を測定した。
・データ処理および分析は、連続測定20個を平均し、空間線量で補正し、
微生物の働きによる放射線量の変化を浮き彫りにるすため、放射能半減期崩壊による減少分を補正し、
さらに直近7日間を移動平均処理することによって放射線特有のデータバラつきを平準化した。
その結果は次のとおり。
①土壌から発するγ放射線は、微生物の働きで低減することが明らかになった
②γ放射線の低減は光合成細菌の働きによるものと思われる
③放射線の低下の割合は、底面よりも土壌表面の方が大きい
④ひとたび微生物が働いた後には、土壌が乾燥しても土壌表面の放射線低減力は残っている
⑤光合成細菌を含む微生物を使って人為的に放射線を軽減することができる
⑥微生物の働きによる「放射能崩壊加速説」の信ぴょう性が高まった。
-----------------------------------------------------------------------------------------
既に「放射能低減実験その2」にてライブ的に経過を報告してきたが、多少の誇張・推測・勘違いもあり、
改めてこの場で実験全体を整理し考察し直した。
なお、実験が未熟だった放射能低減実験その1 も参考まで。
-----------------------------------------------------------------------------------------
1.実験の目的とその背景
(1) 自分の手で土壌微生物が放射能を低減する客観的なデータをつかみ、
その事実詳細を誰にでも分かりやすい形で公表すること
・放射能汚染や風評で苦しんでいる東北・北関東および健康不安を抱いている全国の生活者のみなさんに、
・誰にでも簡単にできる「放射能汚染対策の可能性」を知って欲しく、、
・家族を守るためにも、自宅や地域等にて放射能対策の活動をしていきたいものと願っている。
(2) 家族の健康や食の安全を国や行政だけに任せてはいけない
・チェリノブイリ原発事故のあと、農地にEM(有用微生物群)を施すことによって、
放射能が低減できることが実証実験で報告されていた。
・植物への放射能吸収が抑えられたこと
・農地の放射能が低減したこと
・また、フクシマの高度に放射能汚染された地区で、EMなどで有用発酵した肥料を使った農業では、
・植物に吸収される放射能はND(検出限界外)であったこと
・EMを散布した短期間に土壌の放射能が低減したこと
など、幾多も報告されている。
・しかし、国や行政や多くの御用学者やメディアまでもが、このような微生物による放射能低減効果の
事実を認めようとしないばかりか、微生物を拒絶または忌み嫌う姿勢が続いている。
放射能汚染で苦しむのは、汚染地域居住者や子どもや健康に不安を持つな生活者であり、
農林漁業従事者をはじめ、風評被害の産業である。
国は、除染に莫大なお金を投入しながら、環境回復のための基本的な放射能対策の指導ができないまま、
住民にも国民にも世界に対しても、深刻な事態を軽視し、安全だと言い続けて責任を逃れようとしている。
この日本を、どうしようもない国にしてはならない、生活者が安心して暮らせる国にしていきたものである。
2.実験の準備
放射能低減実験は、平成25年5月連休で実験環境を用意し、5月8日から6月16日まで「実験その1」を行い、
6月26日から実験方法を新たにして今回の「実験その2」が始まった。



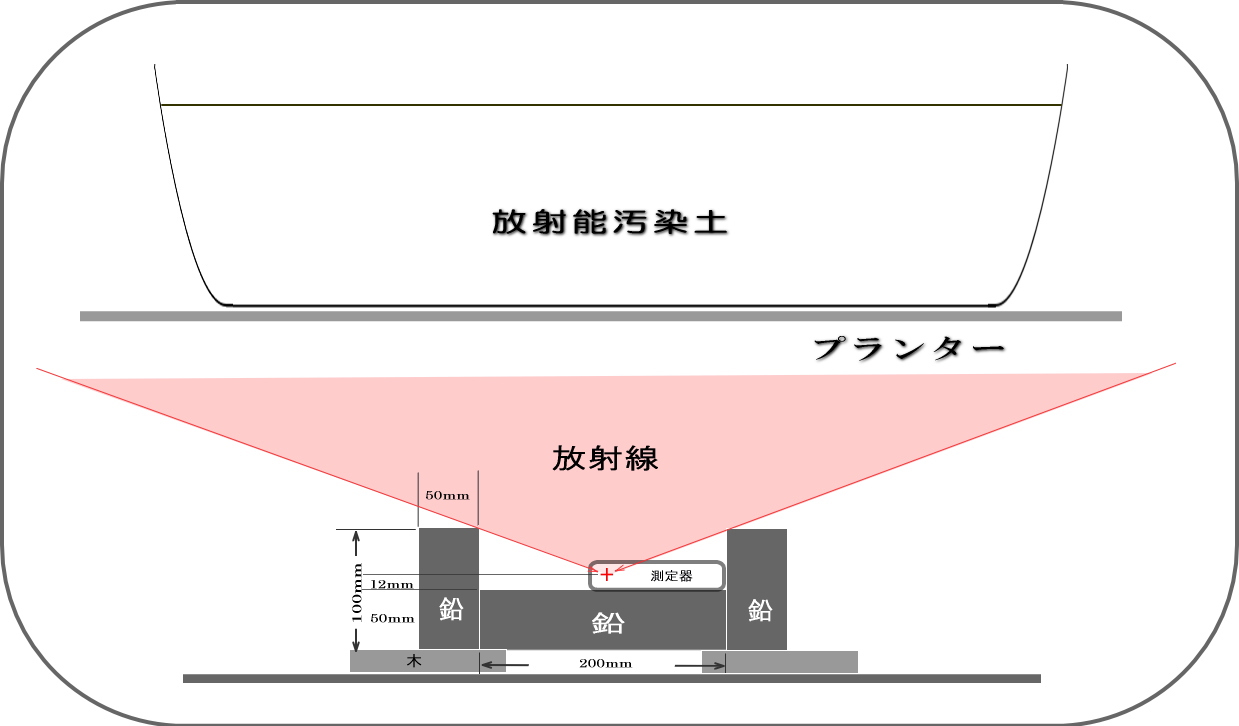

放射能汚染土に混ぜた微生物関連資材や実験経緯などは、別トピック「放射能低減実験その2」 に
ライブ的にUPしてきたのでご覧ください。
3.計測方法とデータ表
(1) 実験の概要
(実験方法) 放射能汚染土に微生物資材を混ぜたプランターにおいて、
土壌表面およびプランター底面から放射される放射線量を計ること
(実験者) 高橋剛(このサイト運営者)
(実験場所) 岩手県紫波郡矢巾町赤林(農村地帯)に設置したミニハウス
(実験期間) 平成25年6月26日~同9月19日(85日間)
(微生物叢環境づくり資材) モミ殻燻炭、ピートモス、培養土、EMボカシ、
EMセラミックパウダー、EM活性液、EM3(光合成細菌強化EM)など
(植えた植物) ヒメイワダレ草
(実験データ) プランター底面とプランター土壌表面の2ヶ所の放射線量を計り、
各々連続する20データを平均した
(求める分析データ) 微生物群などの投入や環境変化に応じた放射線量の変化
(2) Excelによるデータ表は次のとおり
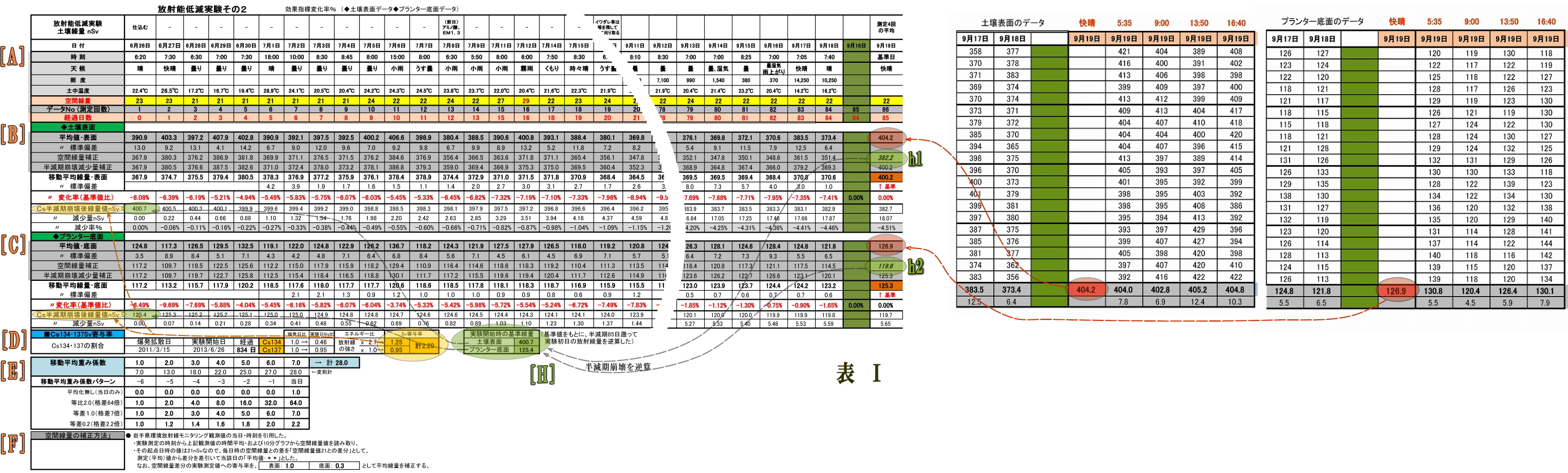
(表の構成)
・左側は、総括表である (中間時期を割愛し、実験初期と実験終期と並べた)
・右側は、別シートに収めた計測データ表の終期だけのもので、
最終日は時間をずらした計測値4回分の平均を求め、それを総括表にリンクしている。
詳しくは、子スレッドにて 「データ分析表の説明」 としてUPしている。
4.実験の経過
(1) 実験は85日間、土壌表面およびプランター底面から発生する放射線量を測定
・データ測定は、ほぼ毎日1回測定し、連続20個のデータを平均化し、それを元に、
・外から飛び込んでく空間線量を補正し、
・さらに汚染土に含まれる放射能自体の物理的半減期崩壊による減少値分を補正することで、
微生物だけの影響による放射線量の変化を知る というものである。
・そして、バラつきある直近7日間のデータを移動平均処理にて平準化した。
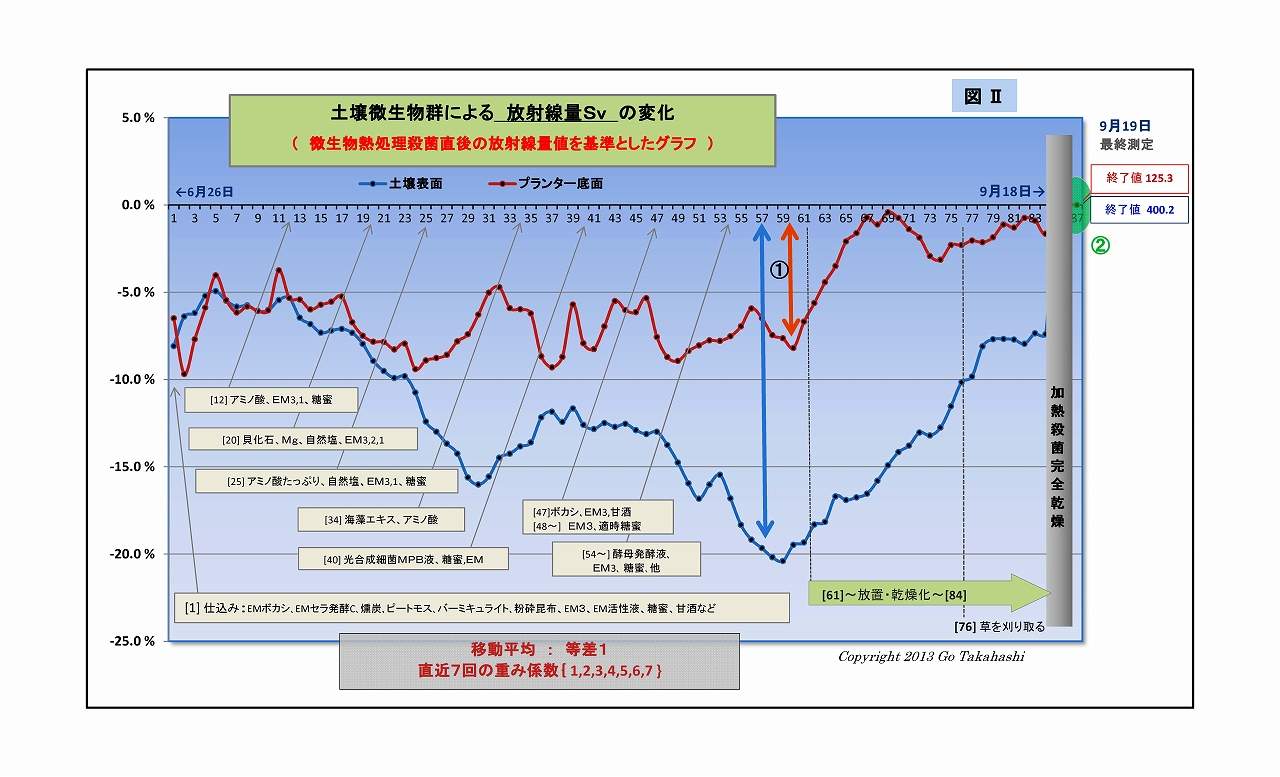
・下図は、このようにして求めた放射線量Svの変化をグラフ化したものである
・変化の基準としたデータは、微生物が含まれない汚染土の放射線量値である。
(実験最終段階で実験土壌を加熱殺菌して測定した)
・グラフの大まかな見方は、
・ヨコ軸は85日間の時間軸、タテ軸は基準値からの変化%である
・図中の①に示す幅の大きさが「微生物群の働きによる放射線の減少量」 である。
・スタート時点で、すでに-6%以上も下がっている理由は、
土壌には「実験その2」以前から含まれている微生物が、すでに働いていること、
そして、この実験開始時に、更に新たな微生物関連資材を加えためである。
・また、グラフは直近7日間の移動平均した値のものなので、放射線独特のバラつきは平準化されているが、
実験直後は直近データが足りないために、初日から数日間は平準化しきれず生データに近いものとなっている。
・なお、グラフの右後半61日目から放射線量が上昇し続けた理由は、
実験終盤の目的の一つである「意図的に土壌の乾燥を促した」したもので、
実験当初の放射線量に比べて増えている理由は、「放射線の減少量(図中の①)が減った」 ということである、
決して「微生物によって放射線量は増えたということではない」 ことを強調しておく。
(2) 実験は、大きく3フェーズになっている
[第一フェーズ] 初日~31日目
・開始から2週間目頃から、放射線量が少しずつ下がってきた。
・確実に下げたいがために、微生物叢を豊かにするためのいろんな資材を適時に散布した。
特に光合成細菌が増えて活性化できる環境づくりを強く意識した
・しかし、25日目から、底面放射線が上昇に反転し、
続く31日目からは、表面放射線量も低下が止まり反転上昇した。
・それまで順調に低下したものが、なぜ反転上昇したのか、分からずに頭を抱えてしまった。
[第二フェーズ] 32日目~60日目
・しばらくは上昇が続いたが、光合成細菌にとって有用と思われる様々な資材を試行錯誤で
散布はしてみたが、放射線量をコントロールしかねていた。
・しかし、これまでは光合成細菌だけに目を向けていたけれど、酵母菌や乳酸菌などの共生菌を
整えてみよう思い、47日目に自家製の甘酒の希釈液など酵母菌を増やす資材を散布したら、
表面および底面の放射線量が再び勢いよく下降しはじめた。
・この頃になると、ヒメイワダレ草は勢いを増してきている。


[第三フェーズ] 61日目~最終85日目
・60日目になると、またもや土壌表面放射線の下降が止まり上昇し始めた。
・これをキッカケに、実験終盤のテーマとしていた「土壌の乾燥(微生物の活性を落す)」 に進んだ結果、
・予想通り、放射線低減量が減りつづけ、平衡に達しようとしていた。
乾燥をもっと加速させるべく草を刈り取ってはみたが、切った茎から新芽が出てくる始末、
プランターであっても環境空気中に湿度がある限り完全に乾燥するのは無理だと悟った。
(なお、これ以降、刈り取った草も含めて測定している)
そして最後の手段、微生物を加熱殺菌処理して、汚染土そのものの放射線量を測定した。
・微生物の働きを完全の停止するには、加熱殺菌が確実であろうと判断し、
台所のハイカロリーコンロで中華鍋で土を焼いて微生物を加熱殺菌し、
(少なくとも微生物の細胞タンパクは破壊し光合成機能は喪失したと思う)
・それをプランターに戻して翌日に最後の測定を行った。
・時間をあけながら4回測定し、
・その平均値を最終日の測定値とする
この値が「微生物に影響を受けない汚染土の放射線量」なので、この値を分析やグラフ化の基準値とした。









5.考察
まず最初に、この実験は、放射線測定器があれば、誰でも容易に試すことができることを強調しておく。
また、分析データは、次のよう処理によって、公開に耐えるに足りる信頼性があるものと思っている。
・1回の測定データ連続23個のうち、最初の3個を捨て、残り20個を平均していること
・平均値に含まれる空間線量(天候によって変動)を補正していること
・更に、日時の違いによってバラつき気味な値を、移動平均処理にて平準化していること
(1) 土壌から発するγ放射線は、微生物の働きで低減することが明らかになった
・グラフ全体から、明らかに微生物の働きによって放射線量が変化することが分かった。
・土壌微生物叢を、外部から人的にコントロールすることで、土壌放射線量を低減できることが証明された。
・従来の放射能に対する科学者たちの常識では、
・γ放射線の透過力はとても強く、金属をも通過するので、
・微生物ごときで放射線は弱まるハズがない!
と、いうことであった思われるが、
これが、いとも簡単に否定されることが実証された(新知見)
(2) γ放射線の低減は光合成細菌の働きによるものと思われる (微生物による放射線吸収説)
・光合成細菌は、光(電磁波)エネルギーを吸収して光合成を行う微生物である。
γ線も電磁波の一種であり、放射線を当てることで光合成細菌は増殖することも分かっている。
・放射能γ線エネルギーは、光合成細菌の細胞内の光合成メカニズムによって利用・吸収されるので、
吸収された分だけγ線エネルギーは減衰し、光合成細菌の密度が高いほど減衰量は大きくなる。
放射線の減衰とは、つまりは 「放射線の低減」 のことである。
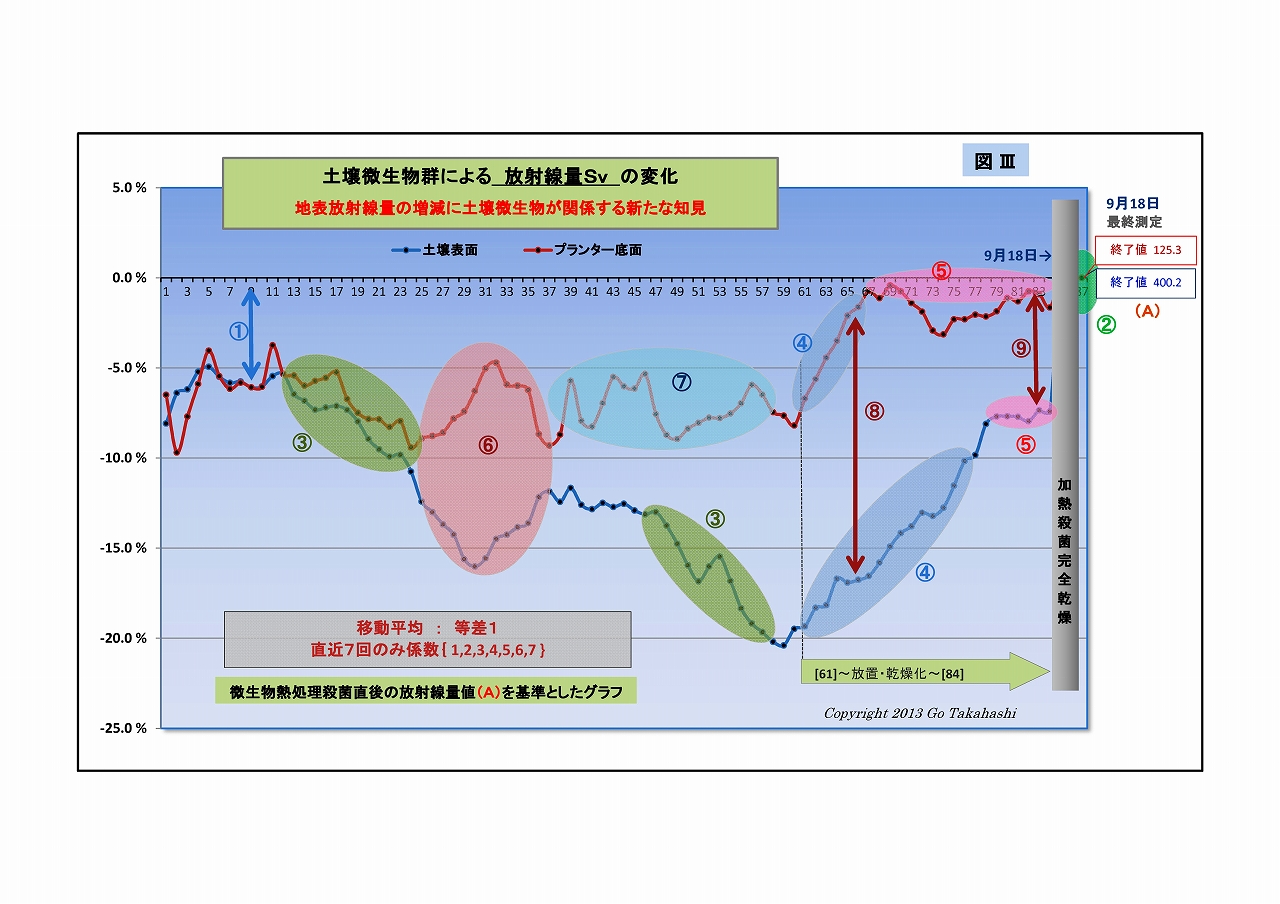
(3) 放射線の低下の割合は、⑧に見られるように、底面よりも土壌表面の方が大きい
・これは、表面側の方が減衰量は大きいことを意味している
光合成細菌は、土壌の深場より表面近くの方が増殖しやすく密度が濃くなっている、と思われる。
・表面と底面の格差は、微生物叢が豊かになっているのだろか、時間経過とともに大きくなっている。
・しかし、④⑤に見られるように、土壌の乾燥が進んでも明確な差は残っている。
(4) 一たび微生物が働いた後には、土壌が乾燥しても土壌表面の放射線低減力は残っている
・乾燥が進んだ④~⑤を見ると、底面び方は微生物の働きがほぼ止まったのに対し、
表面の方は、低減が-7%ほどで止まったままになっているが、
・光合成細菌は土壌が極度の乾燥状態であっても、γ放射線を遮蔽し続ける能力が残っているものと思われる。
(5) 光合成細菌を含む微生物を使って人為的に放射線を軽減することができる
・フェーズ1の③に見られる放射線の低下は、光合成細菌をはじめとする微生物叢を豊かにするための
有効な資材を放射線汚染土に混ぜた結果であること。
・フェーズ2の③に見られる放射線の低下は、更なる低下を目指して、人為的に微生物叢をコントロール
した結果であること。
・この時、特に意図したことは、光合成細菌の働きを助けるために酵母菌や乳酸菌を補強したことである。
このように、人為的にγ放射線を減らすことができることが実証された。
(6) 微生物の働きによる「放射能崩壊加速説」が浮上した
・⑥に着目すると、「変化の方向が表面と底面で逆である」 この原因は何だろうか?
・それまでは表面・底面ともに放射線がムーズに低下したのは放射線吸収説で説明できる、
しかし25日目を境に突然底面だけが上昇方向に転じたが、
土壌表面は依然として下がり続けているので、光合成細菌は増え続けていると考えられるが、
だとすると底面方向も下がっていいものだが、実際の挙動は違っていた。
・考えられる一つは、
光合成細菌には鞭毛があって、光に向かって進む性質があるので土壌表面に移動したのではないか、
という見方である(光合成細菌移動説)。
しかし、⑥の後半などでは、その逆方向の動きも散見されるので、移動説だけの説明には無理がある。
・別の考えもある、
光合成細菌は「放射能崩壊を早める」という説がすでに囁かれていたものだが、
果たして、この現象がそうなのだろうか?
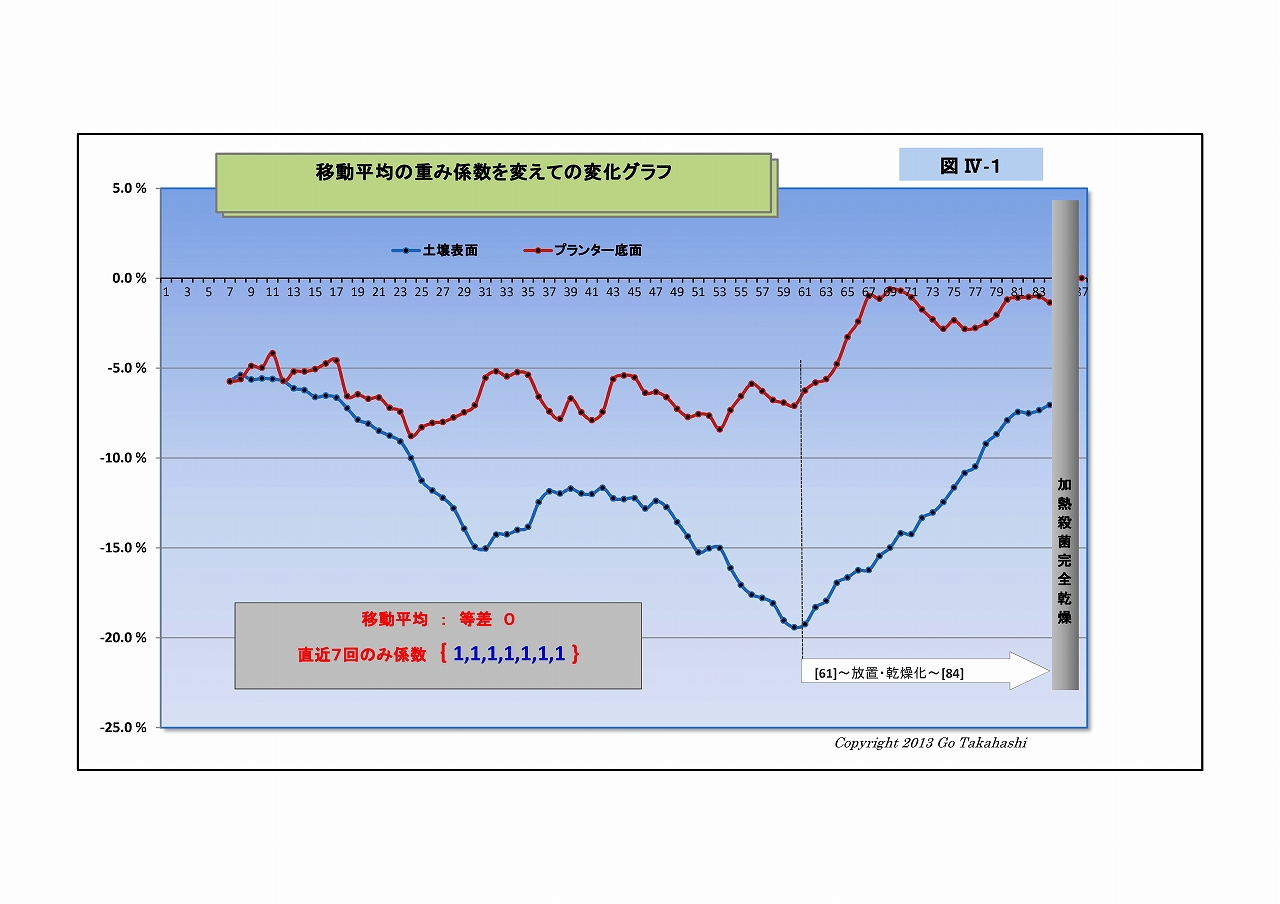
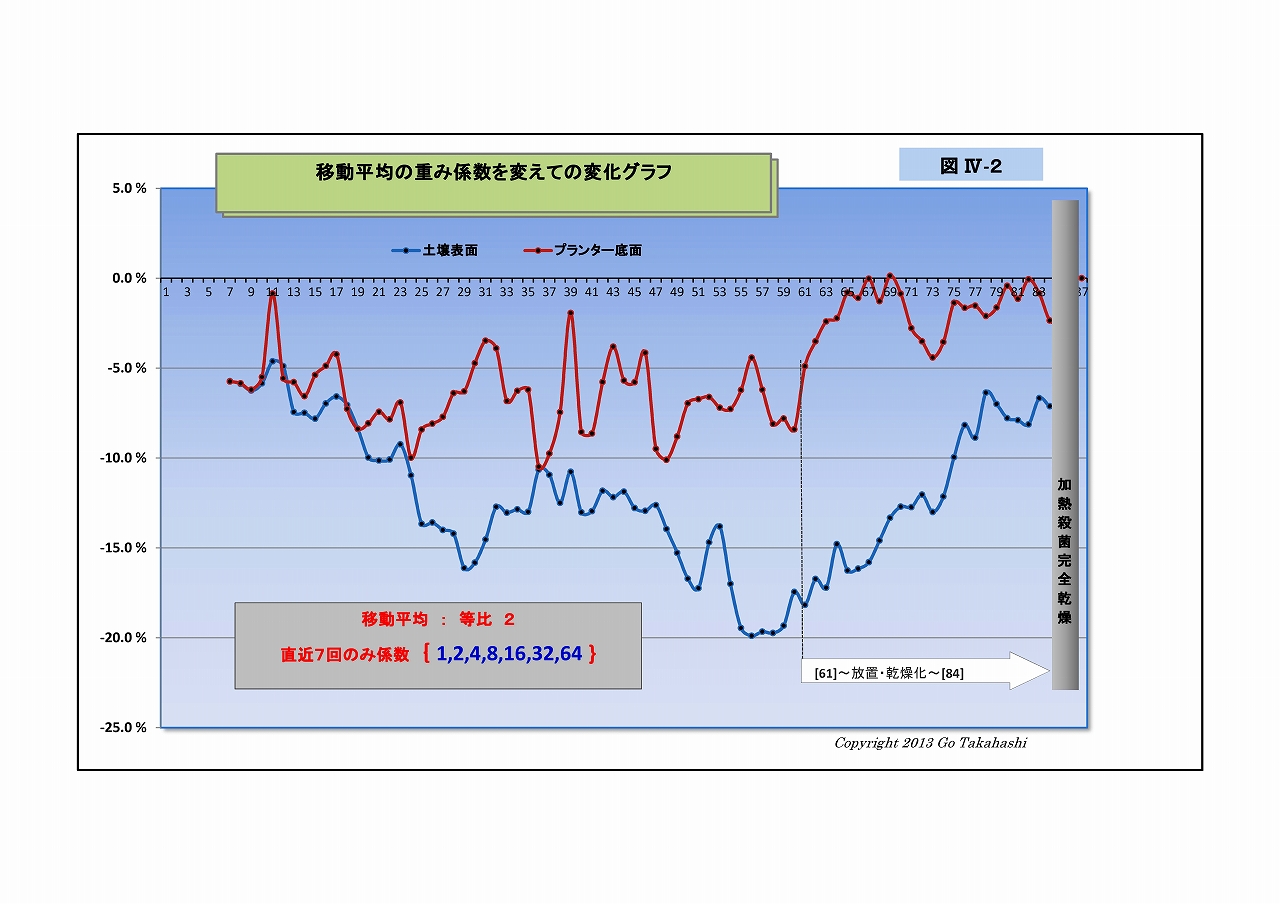
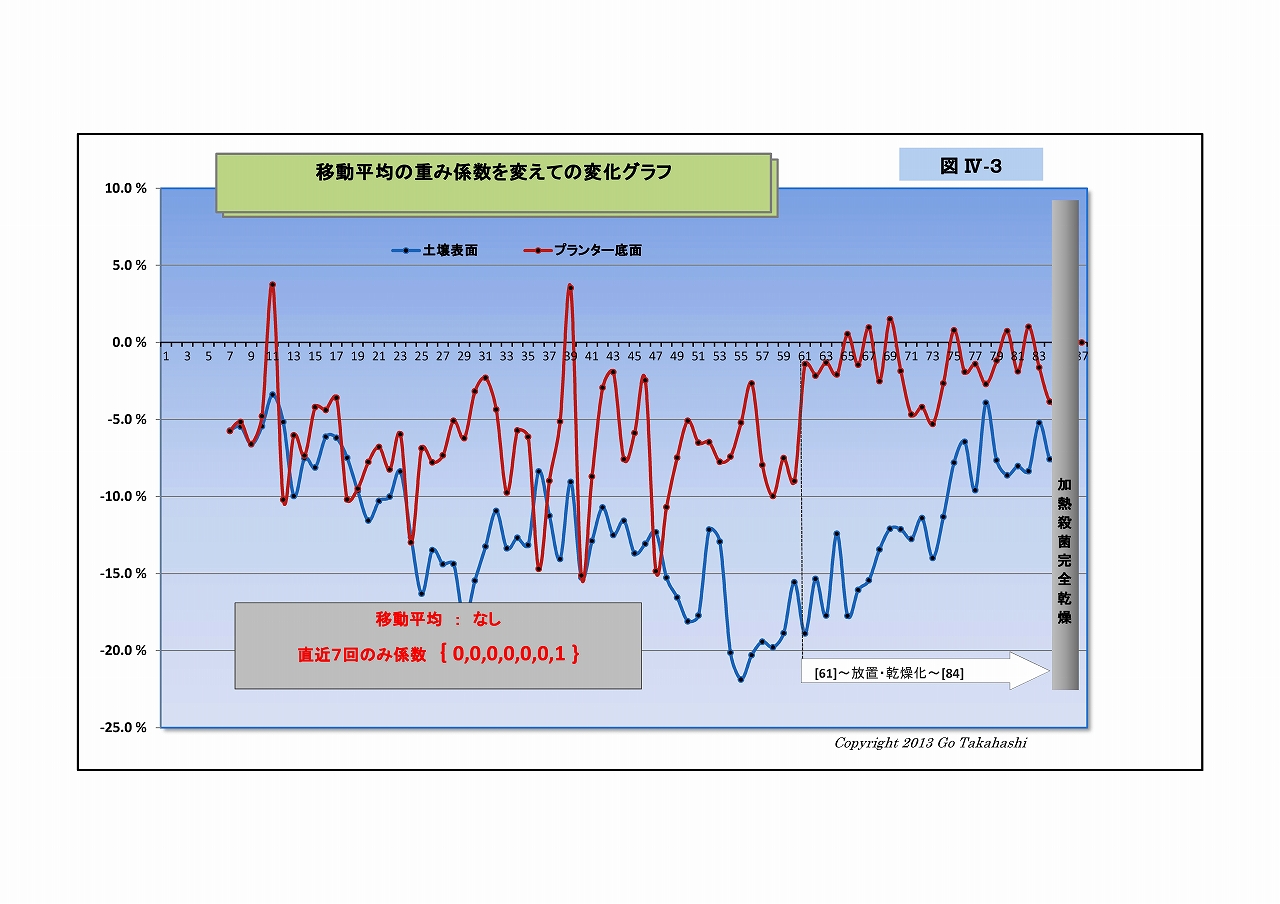
・そこで、移動平均の重み係数をいろいろと変えてグラフにした(下にUP)。
右端グラフ「重み係数なし(生データ)」をよく見ると、
・放射線量は激しく振れているが放射線特有のバラツキとは異なった挙動であること、
・表面と底面方動きの向きが反対になっている箇所が随所にあること、
・時には土固有の放射線(基準値)をオーバーする箇所も散見されること、
・そして⑦を見ると、表面の変動よりも底面の変動の方が激しいことなど、
放射能吸収説や光合成細菌移動説を併しても説明つかない挙動ある。
・ひょっとして、微生物叢の働きによる「放射能吸収」と「放射能崩壊加速=放射線増加」が、
並行して起こり、互いに拮抗している状況のように思えてならない。
・「放射能崩壊加速説」なら、半減期より早く崩壊する分だけけ放射能は低減される、ということだ。
・「放射能崩壊加速説」の裏付けには、実験前後で放射線Bq値が減少している証が必要があるが、
実験その2では、実験前に根拠となるデータを取っていなかったことが悔やまれる。
(すでに実験その3が進行しているので、その実験の終わりに、加熱殺菌して放射線量を計るつもりである)
6.雑感・提言
(1) 土壌の微生物叢を豊かにすることで、人体に悪影響をおよぼす放射線Svを減らすことできる
・自宅の庭や花壇や塀周りや地域の子どもたちのの遊び場など、緑を豊かにしましょう、
・地肌を出さないような管理をしたいものですね。
・放射線量は、10~20%は簡単に減らせると思います。
(2) 誰にでも放射線を低減できる方法
・すでに、福島をはじめ、宮城、栃木、茨城、群馬など各地の市民、ボランティ、農林畜産業の方々が、
自発的にEM(有用微生物群)を活用した射能対策を行って、成果をあげており、
EMの具体的な活用方法と成果事例が多くのサイトにUPされているので、それを参考にしていただきたい。
・しかし、必ずしもEMにこだわる必要はない。
光合成細菌はいろんなメーカから流通されているし、
米ヌカなどを散布して、その上から糖蜜などでしっかり乳酸発酵した培養液の希釈液を散布するだけで
良好な微生物叢が形成され、自然に光合成細菌が増えていくものである。
・私の実験でも、EMだけに頼ることはしていないが、
光合成細菌をはじめ、いかに土壌微生物叢を機能的に豊かにするかに手段は選ばなかった。
(簡単な放射能の低減方法の提案)
・やはり、EM発酵液は必須です、手軽に確実に乳酸菌・酵母菌が培養できるからです。
全国どこでも、お近くに培養して実践しているボランティアな人が必ずいますから、
一度訪ねてみることが早道です。
・そのEM希釈液に光合成細菌を少し足して、散布するだけです。
散布直前に米ヌカなどを薄く撒けば、ベターな微生物叢ができる。
・お庭などでは、土を丸出しにしないことです、
・放射能を含んだ粉塵が肺を直撃しますから、
・ぜひカバープランツを植えましょう、
芝生もいいけど刈込などの管理が面倒でしょうから、
ヒメイワダレ草がおススメです、
・繁殖力旺盛、根は深く張り、背丈は伸びず地面を這って広がる、踏まれても強く、雑草にも強い、
冬は表面は枯れるけど根はしっかりと生きていて、5月ころからは緑の絨毯にる、
・年中小さな花が咲いくれるのもうれしいし・・・。
(3) 研究者・科学者のみなさんにお願いしたい
・いまは、生活や環境活動や農業などに微生物を使った活動や実践は珍しいものではなくなった、
しかし、微生物による放射能低減に関する研究は非常に少ない現状にある。
私の実験のような結果は、誰にでも簡単に再現できるものであり、
再現性ある現象には、後日科学的に証明されてきたことことが科学の歴史でもあった。
・私の実験でも、放射能に及ぼす光合成細菌の働きにはナゾが多いことがわかった。
・放射線吸収し放射線を減衰させること
・放射能の崩壊を即しているのではないか?
・逆に、放射能の崩壊を抑えているのかも知れない?
・放射能核種の元素転換を起こしているのかも知れない?
・・・・・
ぜひ、研究して欲しい、科学的に証明して欲しい、その知見などをネット公開して欲しいと願っています。
(4) どこでも行われている 放射線量Bq測定方法は、果たしては正しい値を出せるのだろうか?
・放射能の強さ計るには、放射線を検知し、線量をカウントしてから、Bq,CPM,Svを導きだしている。
・ところが、私の実験で分かるように、土壌微生物の密度・活性度の違いや、
乾燥の度合、採取する深さの違いなどで、放射線カウント数も変わることが分かった。
・放射線Bqベクレル測定の方法を調べてみると、「土壌サンプルは乾燥すること」とは明記されていないのだ。
・土壌微生物の働きで放射線が弱まる、という認識が全くないいからだと思われてならない。
正しく計るには「土壌を加熱殺菌」すべきである、と考える。
--------------------------------------------------------------------------------------
ぜひ、私の実験方法と導いた結論めいたことなどを、一人でも多くの方々に追試していただき、
より普遍的な放射能対策の知恵として広がってもらいたいと念じています。
go
投稿数: 125
投稿数: 125
以下、データ分析に使ったExcel表のアイテムを説明する。
[全体の構造]
・左側は、総括表である(中間を飛ばし、実験初期と実験終期と並べた)
・右側は、別シートの計測データ表の終期のものであり、
最終日は時間をずらした計測値4回分の平均を求め、総括表に代入している
[A] 基本部
(基本)測定日、測定時刻、データNo(測定回数)、経過日数、天候、土中温度、後半から照度
(空間線量)
・空間線量は天候によって変動するので、計測データに対して補正するためのもの
・この空間線量は、環境放射線モニタリングシステムから「盛岡市」の観測データを利用した。
(この観測点は当実験場所から8キロと最も近い)
[B] 土壌表面
(平均値・表面) 別シートの土壌表面測定データ20回の平均値とリンク
( 〃 標準偏差) 測定データ20回の標準偏差値
( 空間線量補正) 平均値から空間線量を差引く
( 半減期崩壊減少量補正) 計測データには半減期崩壊による減少量が含まれているので、
この減少量を補正(後述の減少量を加算)する。
このことによって、微生物の働きだけによる線量の変化が求めることができる。
(移動平均線量・表面) 直前7日間の移動平均値。
移動平均処理することにより、測定日時によるバラつきが平準化される
・なお7日に満たない実験開始直の6日間は、日数分だけ移動平均する
( 〃 標準偏差) 移動平均区間7日間分の標準偏差
( 〃 変化率(基準値比)) これが最終的に求めたいデータであり(グラフの元になるデータ)、
実験最終日(9/19)の加熱殺菌した直後の放射線量基準値との比率%である。
(Cs半減期崩壊後線量量nSv)
Cs134とCs137が含まれている放射能の半減期崩壊量Svで計算するには単純ではない、
Cs134とCs137とでは、半減期もエネルギーの強さも異なるからだ。
そこで後述[D]の「寄与率」を事前に算出しておき、半減期崩壊後の線量Svを求める
( 〃 減少量) 実験開始時点からの減少量
( 〃 減少率%) その減少率
[C] プランター底面
土壌表面の説明と同じ
[D] Cs134・137Sv寄与比
Cs134とCs137の放射能線量Bqが同量であってもエネルギーの強さが異なるので、
寄与率を求めてSvを計算する必要がある。
(Cs134・137の割合)
・フクシマ爆発のときのCs134とCs13のに構成比が同量であるということなので、
爆発拡散日から834日後の実験開始時点の放射能Bq比を求める、
・さらに、Cs134とCs137の個々のγ線エネルギーの強さは分かっているので、
これにBqを乗算することで、放射線量Svの寄与率が求まる。
・[B][C]の「半減期壊後線量値Sv」は、この「寄与率」と「経過日数」
および「実験開始時の基準線量」(崩壊前の線量)から崩壊後線量を求めている。
・なお、「実験開始時の基準線量」(詳細は[H])は、加熱殺菌した汚染土そのものの
計測値(空間線量補正後)をもとに、半減期85日遡って逆算した。
[E] 移動平均重み係数
・放射能は全くランダムに崩壊する。
1回の測定で20データを記帳し、その平均値を当該日時の値としているが、
それでも、時間でバラつきが残る。
・そこで、前7日間の測定値を移動平均処理することで平準化がはかられる。
(移動平均重み係数)
・用意した重み係数パターンをココに挿入することで、そのパターンのグラフが表示される。
(移動平均重み係数パターン)
・測定した放射線量の変化の子細に分析するためのもの。
・区間の後半に重みを大きくすると、
変化が鋭敏となり、土壌表面とプランター底面の変化の相関などを読み取れるし、
・重み係数の差を狭めると、微生物の働きの変化のようすが読み取れる。
[F] (空間線量の補正方法) 記述の通り
go
投稿数: 125
投稿数: 125
「5.考察」では、だいぶ直観的で大胆なことを書いてしまったが、少し検証してみたいと思う。
◆その前に、「微生物で放射能は低減できない」などとネットで発信している人たちがいる。 その根拠は、
① 放射線は、殺菌作用があるので、細菌は生きていけない。
② 光合成細菌が吸収できる光は可視光線近辺だけである。γ放射線は波長が短か過ぎるので吸収できない。
というものらしい
すごく尤もらしい根拠をあげているようだが、間違いも甚だしいですね。
ぜひ一度、私の実証実験のグラフを見ていただきたい、
明らかに「土壌微生物の働きにより放射線が低減している」ことが分かるはずである。
1.「土壌から発するγ放射線は微生物の働きで低減することが明らかになった」と明言した。
グラフを見れば一目瞭然である。以下、
・データ処理は適正に行われていること
・土壌微生物も働きをコントロールしていること
について、整理しながら説明する。
このグラフは、実験プランターの土壌表面の放射線量の変化を示したものである。
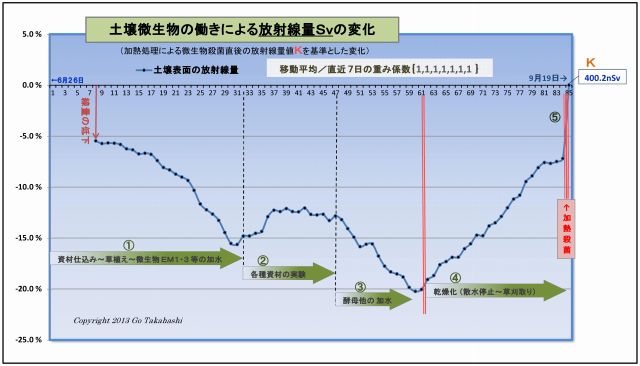
(1) まず最初に、このグラフの元となったデータは適正に処理されている。
・放射線量を連続20点(=200秒)を測定し、それを平均処理し、
・天候等によって変動しがちな空間線量値を岩手県の観測データ値にて補正(減算)し、
・放射能Cs134・Cs137の実験開始からの半減期崩壊値を計算して補正(半減期低下分を加算)し、
・最後に、直前7日間を移動平均して日間のランダム性を平準化した。
このように処理したデータのグラフに見られる放射線量の変化は、
「土壌微生物の働きによるものである」と明言できよう。
(2) 断定できるもう一つの理由「土壌微生物の働きはコントロール」されていることである。
つまり、このデータの変動は偶然に計測さたものではなく、
実験の過程で、微生物の働きを「意図的に操作加減した」結果である、例えば、
・図中①は、実験開始に先立ち、微生物の働きを促進するための微生物と有機資材を汚染土壌と混ぜて、
ヒメイワダレ草も定植したことで光合成細菌をはじめとする土壌微生物叢が豊かになり、
それに伴って、放射線は順調に下がっていること。
・図中②は、放射線の低下が止まり、反転上昇したため、様々な資材(光合成細菌の栄養など)を
試行錯誤的に投入して観察したものであること。
・③は、光合成細菌の働きを良くするために、他の共生微生物を意識的に投入したものである。
例えば「甘酒希釈液」は酵母菌の宝庫であり、この投入で放射線の低下が再び始まり、
③半ばの反転上昇に対しては、「酵母菌増強の発酵液」を散布したこと再び線量は低下した。
この下がった原因は、酵母菌と光合成細菌は共生関係にある、
・光合成細菌がよりいっそう増えるためにはCO2が必要であるが、酵母菌はCO2を排出してくれる。
・一方、酵母菌が増えるためには糖分が必要だが、光合成細菌はブドウ糖を産生する。
・なお、乳酸菌も働くことでphが下がり、腐敗菌が抑えられることで土壌微生物叢が
よりいっそう豊かになる。EM発酵液には乳酸菌などが豊富に含まれている。
・図中④は、資材や水の投入を完全に止め、土壌の乾燥化を促したことによって土壌微生物の働きが弱まり、
その結果として放射線量は上昇(=線量低下が減少)しつづけたこと。
・⑤は、土壌を中華鍋で高熱処理することで水分が皆無となり、
微生物の働きが止ったことで、汚染土壌の本来の放射能によるγ放射線量Kを計測することができ、
このKを基準値として、実験初回からの全データの線量比率を再計算して、上記グラフができあがった。
以上のことから、微生物の働きを意図的にコントロールすることで土壌表面を突き抜けるγ放射線を低減することができた。
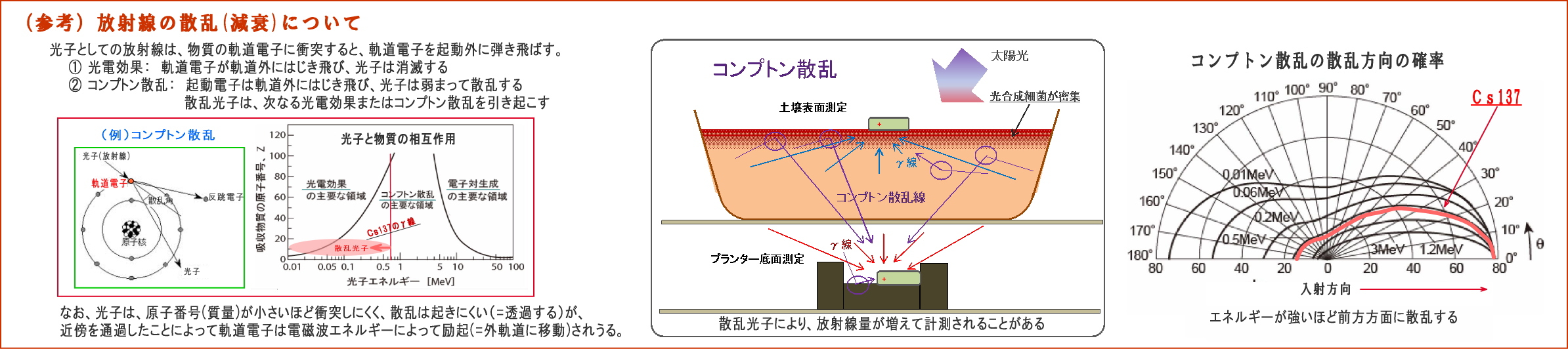
(3) 放射線量の変化は、コンプトン散乱によるものではない
実験による放射線量の変化は、放射線等の散乱の影響を受けているのではないか、との指摘はありそうだ。
① 放射線は、電磁波であるとともに光子(粒子性)でもあるので、
強いエネルギーを持つ光子は、物質に衝突すると相互作用を起こすことがある。 例えば、入射した光子が、
・物質の軌道電子をはじき飛ばして消滅する(光電効果)、或いは、
・軌道電子をはじき飛ばし、自らはエネルギーを弱め、方向を変えて散乱する(コンプトン散乱)
そこで下図に、コンプトン散乱についてまとめてみた。
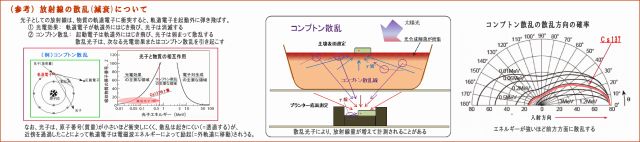
② 入射したγ放射線は、物質との相互作用によって弱められるものであるが、
・しかし、電子番号の低い元素で構成される物質は、ほとんど減衰せずに透過する、
・原子の大きさ(通過断面)に比べて軌道電子の密度が小さいので、光子(粒子)との衝突の確率が少ない。
(原子番号は原子核の陽子の数であり、軌道電子の数でもある)
・微生物を構成する脂質やンパク質は、大半が原子番号の低いNCHOなどで構成されているので、
放射線は軌道電子にほとんど衝突せず、通過することが多い。
③ 今回の実験期間中で変化した物質は、有機資材の投入、土中の草の根の増加だけだが、
これらの増加によって放射線の相互作用が高まり、線量が変化するのだろうか?
・そこで気になるのは、23日目に少量の貝化石を土壌表面にパラパラと撒いたことだが、
その直後の土壌表面線量は、特に目立った変化はなかったので、
コンプトン散乱反射波や光電効果の影響はほとんど無かったものと言えよう。
・他の投入資材は水溶性であるので、プランターの上下全体に行き渡っている。
・草の根は、期間後半は底面に毛根が密集してきけれど、底面線量がそれによって減衰した形跡はみあたらない。
・いずれにしても、光電効果やコンプトン散乱が多少は起こったのだろうけど、
グラフの線量変化は、微生物の働きによって引き起こされたものであることは明らかである。
以上のことから、金属をも透過するほどのエネルギーの強いγ線であっても、
・土壌微生物は放射線を吸収している
・微生物の量と活性度合いで吸収量は増加する
という結論に行きつく。
なお参考までに、フクイチ原発事故以来、福島県はもとより北関東や宮城県などにおいて、
EMを定期的に散布することで「放射線量が下がる」ことが、フォーラムやネットなどで多く報告されているが、
私の実証実験の結果からみても、納得できるものであることを付記しておきたい。
2.「γ放射線の低減は光合成細菌の働きによるものと思われる」
これは、次の根拠によるものである。
・放射線量は、土壌表面の方がプランター底面よりも低下(=吸収)が大きいこと。
・光合成細菌は、太陽光の当たる土壌表面に密集すること。
・光子エネルギーを利用吸収できる微生物は光合成細菌だけであること。
(1) γ放射線の物質の透過力は非常に強い
・Cs137のγ放射線は、可視光線と同じ光子ではあるが、可視光線より遥かに強い(30万倍)エネルギーを持っている。
・今回の実験では、土壌表面では、最大20%ほど放射線量が低下しているが、
この低下の量は、厚さ2mmの鉛板で遮蔽した時の減衰量に匹敵する。
・期間を通して、土壌表面と測定器を遮るものは特にない。
(厳密に言えば、草はあるけど、測定器の検出部は草の茎のない処に置いているし、
試しに草の茎や葉を少し寄せ集めてその上から計ってみたが測定データは変わらなかった)
(2) 土壌表面のγ線量の低下は光合成細菌の仕業と思われる
① 光合成細菌の密度は土壌表面の方が遥かに高い
・光合成細菌は、太陽光からエネルギーを受けやすい表面近くに密集(=増殖)する。
このことにより、徐々にではあるが、土中(底面方向)より遥かに密度が高くなっていった。
・実験開始直後のしばらくは、プランター底面も土壌表面も低下率は同じであったが、
微生物叢が活発になりはじめると、次第に土壌表面の方が低下率が大きくなったが、その理由については、
光合成細菌が土壌表面に多く集まってきて、放射線を底面よりも多く吸収していると考える。
(3) 鉛板2mmに相当する放射線の減衰は、微生物タンパクなる単なる物質による減衰であるとは到底考え難い。
・物質による減衰ではなく、「生命体として積極的に吸収している現象」と考えたい、
「吸収」とい表現よりも、「エネルギーを積極的に取り込んでいる」と言った方が正確ではないだろうか。
・プランター内の生体は土壌微生物だけであり、光子エネルギーを利用できるのは生体は光合成細菌しかない。
消去法ではあるが、放射線の光子エネルギーを吸収しているのは光合成細菌である、と断言したい。
光合成細菌が、その密度を高め、活動が活性化するほど、放射線の吸収量は増加する、ということである。
しかし、この「光合成細菌の放射線吸収説」を裏付けるには、難儀するねぇ・・・、
ネットのどこを探しても「光合成細菌はγ放射線を吸収する」という情報は見当たらないのだ
3. 光合成細菌による光合成のメカニズム
そこで、光合成細菌がγ線を吸収できるという根拠を探るため、光合成のメカニズムをまとめてみた。
結論を先に言うなら、
・光合成細菌の基質(=エサ)は、光(光子エネルギー)と二酸化炭素である、これを原料として糖を合成する。
・γ放射線は、可視光線よりも遥かに強い光子エネルギーもっている。
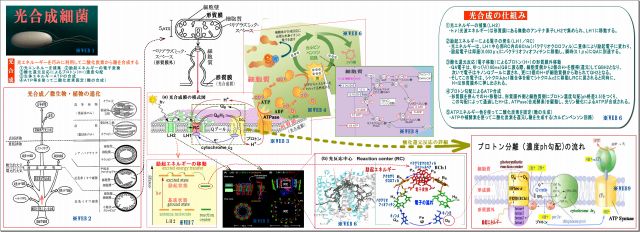
*WEB1 *WEB2 *WEB3 *WEB4 *WEB5 *WEB6 *WEB7 *WEB8 *WEB9
(1) 光合成細菌の細胞膜(形質膜:光合成膜とも言う)には光合成機能が詰まっている
・細胞膜が細胞内に向って小胞上のくびれ(クロマトフォア)がたくさんある。
この図では紅色イオウ細菌のものだが、紅色非イオウ細菌の場合は、クロマトフォアが
細胞質内に更にツリー状に広がっている。
・そのクロマトフォアには、たくさんの「光合成機能セット」が詰まっている、図はその一セットを拡大したもので、
特に光合成膜の面積大半を占めるのは、光エネルギー取り込むアンテナ素子LH2である。
(2) 光合成とは、光エネルギーを巧みに利用して二酸化炭素から糖を合成すること。 その流れは、
① 光エネルギーを捕集
・光で励起された光エネルギーをできるだけ広く集めるためのアンテナ素子がLH2であり、
・その励起エネルギーはLH1の反応中心RC(Reaction Center)に向って伝達される。
・励起エネルギーの伝搬方法は、励起とは軌道電子がより高い軌道に移動することであるが、
その電子は、瞬時に元の軌道に戻る際に他の原子の軌道電子を励起するという。
・この励起エネルギーはLH2→LH1→RCに瞬時に伝搬するらしい。
② 励起エネルギーの電子変換
・励起エネルギーはRCの中核BChl(バクテリオクロロフィル)にて電子に変換され、
バクテリオフィオフィチン→キノンQAへと瞬時に伝達される
③ 酸化還元反応によるプロトン(H+)濃度勾配
・QA電子はQBおよびキノンQプールに移動する際、細胞質側からプロトンH+を取ってQH2となり、
・電子は、後でシトクロム複合体を経てシトクロムc2からRCのヘムに戻されるが、
その前に、H+は形質膜外に押し出され、形質膜の内外にプロトン濃度(ph)勾配ができる。
④ 生体エネルギーATPの合成
・このプロトン濃度(ph)勾配によって、H+はATPase(合成酵素)を通過することで駆動され、
細胞質側で光リン酸化によってATPが合成される。
⑤ ATPを使って二酸化炭素固定(糖の合成)
・細胞質内では、ATPや補酵素を使って二酸化炭素を還元し、糖を合成する(カルビンベンソン回路)
3.光合成細菌が吸収できるのは可視光線だけなのか?
そんなことはない。
・いまや光合成細菌の吸収スペクトルは、研究され公表されているが、
この吸収スペクトルは、可視光線を照射して特定した「光学的スペクトル」のことであり、
光子エネルギー全般について特定されたものではない。
・光合成細菌は、光子エネルギーを吸収するメカニズムが備わっており、
γ放射線やX線などの光子エネルギーをも利用することができる。
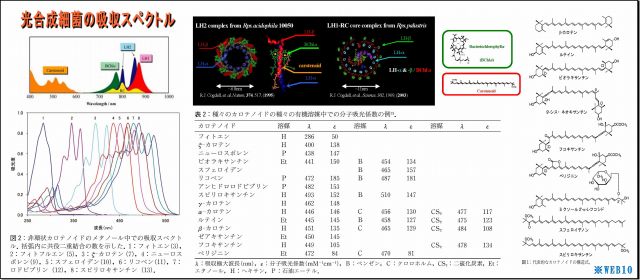
*WEB10
(1) LH2の立体構造を見ると、ヘリカル(螺旋)構造が目についてくる
・これは光も電磁波なので、光を効率よく集めるためなのだろうか、実に見事な形状である、
・また、このLH2の直径は70Å(オングストローム、1Å=0.1nm)=7nm程度であり、ヘリカル径はもっと小さいことから、
可視光線波長(390~750nm)向けアンテナ形状にしては小さいような気もする。
・そもそも原子の大きさは数nmなので、波長と共振できる形状だけから言及するなら、
分子レベルの大きさと構造(すなわち原子配列・形状・光入射向き)や色素複合体などとの関係が重要なのだろう。
・図の「吸収スペクトル」をみると、
・分子構造が複雑で大きいほど赤外線などの長波長に対応しているようであり、
・集光エネルギー終着点であるBChl(バクテリオクロロフィル)もスペクトルを持つが、
これはもっぱら光励起エネルギーの電子変換が主な役割らしい。
・またこれに絡み付くように繋がっているカロチノイドは短波長に対応するという、
・カロチノイドの基幹である共益二重結合の数Nが少ないほど短波長を吸収するようで、
N=3のフィトンでも280nm付近(UV-B中紫外線)止まりのようだ。
以上のことから、
吸収(エネルギー利用)できる光の範囲は、近紫外線~近赤外線(波長300~1,000nm)であるとされており、、
波長0.00187nmであるCs137のγ放射線は、吸収スペクトルから大きく外れていることになる。
このことをもって、「光合成細菌は放射線を吸収できない」「利用されない」と断言する人もいるようだ、
が、果たして、そう断言できるのか?
(2) しかし、私の実験では「γ放射線は光合成細菌によって吸収されている」という実証データがある。
① 公表されたスペクトル論に異を唱えるつもりは毛頭ない。
しかし、もっと短い波長の光子は吸収できない、とは書かれていない。
(3) 研究として公表されている「吸収スペクトル」の特定方法を見ると、
① タンパク質を溶媒に抽出し、近赤外線~可視光線~近紫外線の光を当てて調べたものである。
・つまり、単なる「光学的特性」の結果でしかない、ということだ。
・光は電磁波でもあるので、吸収スペクトルとは、本来電子回路系をも含めた周波数特性のことである。
電子回路でいうLCR(Lコイル、Cコンデンサ、R抵抗)のインピーダンスと周波数との共振性であり、
アンテナとは密接に関係するけど、決してアンテナ素子の物質・形状だけで決まるものではない。
・しかも、電子回路としてみた場合、一連の光化学プロセスは実に厄介な構造であり、いまだ解明されない処も
随所にあることから、周波数スペクトルを特定するには、実際に電子を流して検証する必要がある。
しかし、それは一切なされていない。
②更には、実際にX線や放射線を当てて吸収スペクトルを調べた訳ではもちろんない。
・地球に降り注ぐ太陽エネルギーの大半は可視光線とその前後だから対象外なのだろう。
(参考までに)いまの太陽光線中の割合は、およそ、
遠紫外線1%以下、近紫外線8%程度、可視光線40%、近赤外線20%、中赤外線25%、遠赤外線6% らしい。
・それとも、放射線の強い透過力では、吸収の変化は計測不能に近いからなのだろうか・・・。
・なお、LHCのヘリカル構造の直径は数Åであり、何やらX線帯近くまで対応できているような気がするが。
③そもそも、アンテナ素子LHC(Lh2、Lh1)は、光子エネルギーを受けて電子励起するものである。
・光子エネルギーは何も可視光に限らない、遠紫外線~X線~放射線のエネルギーは、
可視光の数百倍~数万倍~従十万倍もの強い光子エネルギーを持っているものなので、
放射線がLHCを通過する(軌道電子と衝突する確率は低い)だけで容易に軌道電子は励起されよう。
・つまり、アンテナ素子の元素配列や形状(光学的スペクトル)とは無関係に励起される。
・太古の地球では地表に降り注ぐ太陽エネルギーは陽子線・中性子線、X線や放射線が多かった事からして、
その時代に出現した光合成細菌は、生いたち初期からエネルギーの強い放射線やX線を利用していたと思われる。
LHCのヘリカル構造や分子の元素構造や配列形状は、放射線やXが少なくなる時期と合わせて進化したのだろう、
実に巧みに設計されたものと思われる。
④ 放射線の照射で細菌は死滅するということへの反論
・エネルギーMeV以上の強い放射線を局所集中的に照射するなら、死滅すると思われる。
・LHC構造のカロチノイドには、強い照射に対する防御機能(活性酸素を打ち消す役割)もあるらしい
・私の実験では、光合成細菌や酵母などの共生菌が死滅または増殖が阻害されているとは考えられない、
むしろ元気なくらいに感じられる。
・チェルノブイリ事故周辺やフクシマ現場周辺の写真を見る限り、植物が赤茶けて枯れた様子は全く見られない。
・光合成の備わった生物は、放射能に強く、光合成も盛んになるもの、と思われる。
⑤ なお、X線や放射線の吸収性(スペクトル)を確かめたいならば、
・細菌が死んだ状態では計れない、単に透過するだけだから、生きてシステムが働いている状態で測定しなければならない。
・なお、例え細菌が生きていたとしても、負荷が必要である、負荷とは出力負荷=消費のことだ、
つまり、合成されたATPを消費してあげないと、飽和して励起エネルギーの流れはすぐに停滞してしまう、
・電気実験だって、いくら電圧をかけても電力が消費されなければ電流は流れない。
・ATPが消費され、合成された糖などが共生菌によって消費され、更に植物草に消費される、など、
一連の微生物叢や植物との共生が出来上がって、はじめて放射線の吸収が確認できるものと考えたい。
この観点から、私の実験では、詳細なスペクトルまでは調べることはできないが、
間違いなく「放射線を吸収している」ことがを実証しているのである。
4. 光合成細菌の放射線吸収の謎にせまる
光合成細菌は、太古の地球が太陽風(陽子線・放射線・紫外線に晒されていた環境で出現し、進化してきた微生物なので、
放射線に強いというよりも「放射線を利用できるメカニズム」が備わっているものと推測する。
① 光合成細菌に光エネルギー捕集と電子変換の見事なメカニズム
・実に見事に設計されているものと感嘆する。
(イ) 電磁波エネルギーと光子エネルギーの両方を利用できるメカニズムとなっている
(ロ) 電磁波エネルギーは、広く配置したアンテナ素子LHCで捕集する機能、
可能な限り広範囲な可視光線帯のエネルギーを吸収しやすいために
クロロフィルや多様な分子構造をもつカロチノイドの構造と配置によるスペクトルの形成
(ハ) 可視光線より遥かに波長の短い光子(遠紫外線・X線・γ線)のエネルギーは、
光学的スペクトルとは無関係に、直接LHCを励起する。
(ニ) 可視光線近辺の光捕集による励起エネルギーも、より短波長の光子による励起エネルギーも、
バクテリオクロロフィルPにて同一視して電子変換されてる
(ホ) 電子のキノンと結びつき、ATP合成のためのプロトン勾配を作る
(ヘ) 生体エネルギーATPを利用して糖を合成する
・実に見事としか言いようがない。
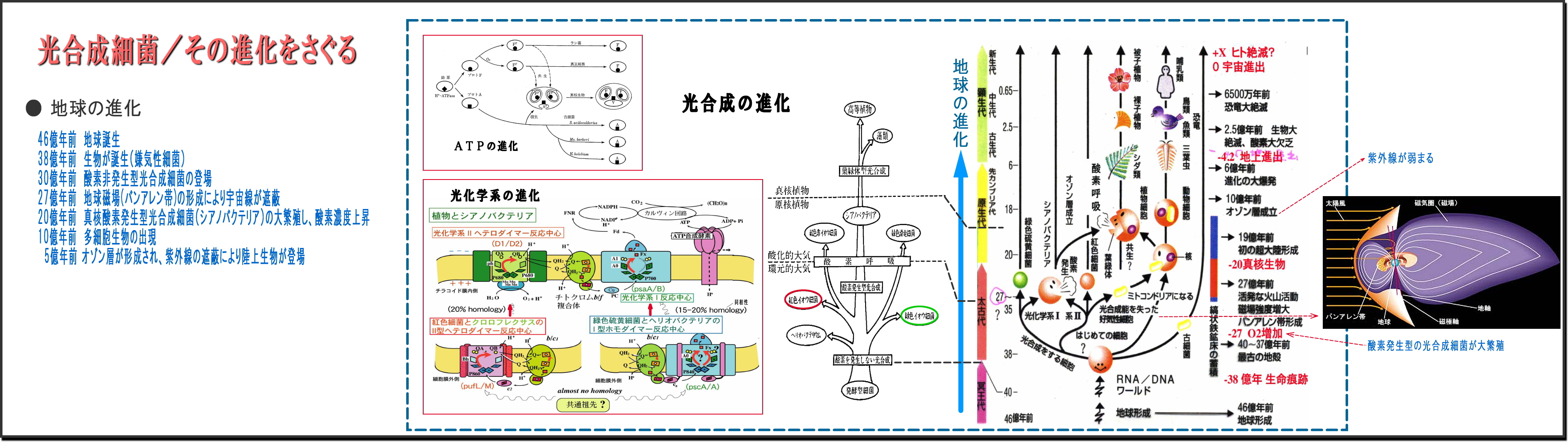
② 地球の進化は生物の進化と共にある
・地球という惑星が誕生したのは46億年前で、最初の生命体誕生は、今なお謎らしい。
38億年前の岩石には、すでに生命の痕跡が見つかっている。
・最初の生物は微生物だが、他から栄養をもらって増殖する従属栄養微生物が多いなか、
自ら栄養を作り出せる独立栄養微生物は光合成細菌だけである、
・この光合成細菌の誕生(微生物の進化)によって、地球は飛躍的に生命力が増してきた。
・光合成細菌の最初は、酸素を発生しない光合成細菌であった。
有毒ガスである硫化水素の多い地球だから、H2Sを還元して光化学する紅色(や緑色)硫黄細菌であり、
次第にH2Oを還元し酸素を排出する非硫黄細菌へと進化する。
・この光合成細菌の光合成機能を受け継いだ原核植物シアノバクテリア(藍藻)へと進化し、
ついには植物の葉のなかの葉緑体へと発展していく。
・この進化のようすはクロマトフォアの密集度合いの推移にみることができる。
・なお余談だが、光合成の中間産物である生体エネルギーATP機能は、動物細胞のミトコンドリア
として細胞内に組込まれていく。
③ 太古の地球には酸素はほとんど無く、やがて活発なマグマが落ち着くころになると、
・地殻が安定し、自転による地磁気が明確になってバンアレン帯が形成される27億年前ころになると、
陽子線や放射線などが含まれる有害な地球への太陽風は次第に弱まり、
やがて酸素の層が厚みを帯びてきて、10億年前頃にはオゾン層ができると紫外線も弱まり、
太陽エネルギーは可視光線がメインとなってきた。
④ 光合成細菌は、このバンアレン帯が出来上がる以前に登場し進化を続けてきた
・バンアレン帯以前の地球は、陽子線や放射線など生体に有害エネルギーが降り注ぐ状態である、
こんな環境でも、光合成細菌のタンパク質などは破壊されないで進化・増殖できたのには、
放射線をも基質(増殖に必要なエサ)として利用してきたと想像する。
・可視光線の数十万倍ものエネルギーのある放射線を、生命の増殖に使わない手はないし、
過酷な環境に順応し、逆に過酷な環境を好むように光合成細菌は進化してきたものと素直に考えたい。
⑤ 光合成細菌の進化について
・上記①に述べたように、光合成の見事なまでの光化学メカニズムは、いきなり完成したわけではない。
・複雑な光学的スペクトルが形成できたのは、進化のずっと後のことで、放射線よりも可視光線の割合が
多くなってきたことに適応しながらの進化ではなかろうか。
・つまり、地球太古の時代は、可視光線よりも遥かにエネルギー高いX線やγ線が降り注いでいたので、
簡単に励起エネルギーを得ることができるシンプルな光化学系メカニズムが最初にできあがった、と考えたい。
そして、バンアレン帯以降あたりから可視光線をも吸収しやすいような光学スペクトル分子構造へと進化してきた。
以上、私がいろいろ調べてみたことを、私の推測も交えて、備忘録的に取りまとめてはみた。
だいぶ謎の核心に近づいてきたのではないかと思っているが、如何なものだろうか
◆その前に、「微生物で放射能は低減できない」などとネットで発信している人たちがいる。 その根拠は、
① 放射線は、殺菌作用があるので、細菌は生きていけない。
② 光合成細菌が吸収できる光は可視光線近辺だけである。γ放射線は波長が短か過ぎるので吸収できない。
というものらしい
すごく尤もらしい根拠をあげているようだが、間違いも甚だしいですね。
ぜひ一度、私の実証実験のグラフを見ていただきたい、
明らかに「土壌微生物の働きにより放射線が低減している」ことが分かるはずである。
1.「土壌から発するγ放射線は微生物の働きで低減することが明らかになった」と明言した。
グラフを見れば一目瞭然である。以下、
・データ処理は適正に行われていること
・土壌微生物も働きをコントロールしていること
について、整理しながら説明する。
このグラフは、実験プランターの土壌表面の放射線量の変化を示したものである。
(1) まず最初に、このグラフの元となったデータは適正に処理されている。
・放射線量を連続20点(=200秒)を測定し、それを平均処理し、
・天候等によって変動しがちな空間線量値を岩手県の観測データ値にて補正(減算)し、
・放射能Cs134・Cs137の実験開始からの半減期崩壊値を計算して補正(半減期低下分を加算)し、
・最後に、直前7日間を移動平均して日間のランダム性を平準化した。
このように処理したデータのグラフに見られる放射線量の変化は、
「土壌微生物の働きによるものである」と明言できよう。
(2) 断定できるもう一つの理由「土壌微生物の働きはコントロール」されていることである。
つまり、このデータの変動は偶然に計測さたものではなく、
実験の過程で、微生物の働きを「意図的に操作加減した」結果である、例えば、
・図中①は、実験開始に先立ち、微生物の働きを促進するための微生物と有機資材を汚染土壌と混ぜて、
ヒメイワダレ草も定植したことで光合成細菌をはじめとする土壌微生物叢が豊かになり、
それに伴って、放射線は順調に下がっていること。
・図中②は、放射線の低下が止まり、反転上昇したため、様々な資材(光合成細菌の栄養など)を
試行錯誤的に投入して観察したものであること。
・③は、光合成細菌の働きを良くするために、他の共生微生物を意識的に投入したものである。
例えば「甘酒希釈液」は酵母菌の宝庫であり、この投入で放射線の低下が再び始まり、
③半ばの反転上昇に対しては、「酵母菌増強の発酵液」を散布したこと再び線量は低下した。
この下がった原因は、酵母菌と光合成細菌は共生関係にある、
・光合成細菌がよりいっそう増えるためにはCO2が必要であるが、酵母菌はCO2を排出してくれる。
・一方、酵母菌が増えるためには糖分が必要だが、光合成細菌はブドウ糖を産生する。
・なお、乳酸菌も働くことでphが下がり、腐敗菌が抑えられることで土壌微生物叢が
よりいっそう豊かになる。EM発酵液には乳酸菌などが豊富に含まれている。
・図中④は、資材や水の投入を完全に止め、土壌の乾燥化を促したことによって土壌微生物の働きが弱まり、
その結果として放射線量は上昇(=線量低下が減少)しつづけたこと。
・⑤は、土壌を中華鍋で高熱処理することで水分が皆無となり、
微生物の働きが止ったことで、汚染土壌の本来の放射能によるγ放射線量Kを計測することができ、
このKを基準値として、実験初回からの全データの線量比率を再計算して、上記グラフができあがった。
以上のことから、微生物の働きを意図的にコントロールすることで土壌表面を突き抜けるγ放射線を低減することができた。
(3) 放射線量の変化は、コンプトン散乱によるものではない
実験による放射線量の変化は、放射線等の散乱の影響を受けているのではないか、との指摘はありそうだ。
① 放射線は、電磁波であるとともに光子(粒子性)でもあるので、
強いエネルギーを持つ光子は、物質に衝突すると相互作用を起こすことがある。 例えば、入射した光子が、
・物質の軌道電子をはじき飛ばして消滅する(光電効果)、或いは、
・軌道電子をはじき飛ばし、自らはエネルギーを弱め、方向を変えて散乱する(コンプトン散乱)
そこで下図に、コンプトン散乱についてまとめてみた。
② 入射したγ放射線は、物質との相互作用によって弱められるものであるが、
・しかし、電子番号の低い元素で構成される物質は、ほとんど減衰せずに透過する、
・原子の大きさ(通過断面)に比べて軌道電子の密度が小さいので、光子(粒子)との衝突の確率が少ない。
(原子番号は原子核の陽子の数であり、軌道電子の数でもある)
・微生物を構成する脂質やンパク質は、大半が原子番号の低いNCHOなどで構成されているので、
放射線は軌道電子にほとんど衝突せず、通過することが多い。
③ 今回の実験期間中で変化した物質は、有機資材の投入、土中の草の根の増加だけだが、
これらの増加によって放射線の相互作用が高まり、線量が変化するのだろうか?
・そこで気になるのは、23日目に少量の貝化石を土壌表面にパラパラと撒いたことだが、
その直後の土壌表面線量は、特に目立った変化はなかったので、
コンプトン散乱反射波や光電効果の影響はほとんど無かったものと言えよう。
・他の投入資材は水溶性であるので、プランターの上下全体に行き渡っている。
・草の根は、期間後半は底面に毛根が密集してきけれど、底面線量がそれによって減衰した形跡はみあたらない。
・いずれにしても、光電効果やコンプトン散乱が多少は起こったのだろうけど、
グラフの線量変化は、微生物の働きによって引き起こされたものであることは明らかである。
以上のことから、金属をも透過するほどのエネルギーの強いγ線であっても、
・土壌微生物は放射線を吸収している
・微生物の量と活性度合いで吸収量は増加する
という結論に行きつく。
なお参考までに、フクイチ原発事故以来、福島県はもとより北関東や宮城県などにおいて、
EMを定期的に散布することで「放射線量が下がる」ことが、フォーラムやネットなどで多く報告されているが、
私の実証実験の結果からみても、納得できるものであることを付記しておきたい。
2.「γ放射線の低減は光合成細菌の働きによるものと思われる」
これは、次の根拠によるものである。
・放射線量は、土壌表面の方がプランター底面よりも低下(=吸収)が大きいこと。
・光合成細菌は、太陽光の当たる土壌表面に密集すること。
・光子エネルギーを利用吸収できる微生物は光合成細菌だけであること。
(1) γ放射線の物質の透過力は非常に強い
・Cs137のγ放射線は、可視光線と同じ光子ではあるが、可視光線より遥かに強い(30万倍)エネルギーを持っている。
・今回の実験では、土壌表面では、最大20%ほど放射線量が低下しているが、
この低下の量は、厚さ2mmの鉛板で遮蔽した時の減衰量に匹敵する。
・期間を通して、土壌表面と測定器を遮るものは特にない。
(厳密に言えば、草はあるけど、測定器の検出部は草の茎のない処に置いているし、
試しに草の茎や葉を少し寄せ集めてその上から計ってみたが測定データは変わらなかった)
(2) 土壌表面のγ線量の低下は光合成細菌の仕業と思われる
① 光合成細菌の密度は土壌表面の方が遥かに高い
・光合成細菌は、太陽光からエネルギーを受けやすい表面近くに密集(=増殖)する。
このことにより、徐々にではあるが、土中(底面方向)より遥かに密度が高くなっていった。
・実験開始直後のしばらくは、プランター底面も土壌表面も低下率は同じであったが、
微生物叢が活発になりはじめると、次第に土壌表面の方が低下率が大きくなったが、その理由については、
光合成細菌が土壌表面に多く集まってきて、放射線を底面よりも多く吸収していると考える。
(3) 鉛板2mmに相当する放射線の減衰は、微生物タンパクなる単なる物質による減衰であるとは到底考え難い。
・物質による減衰ではなく、「生命体として積極的に吸収している現象」と考えたい、
「吸収」とい表現よりも、「エネルギーを積極的に取り込んでいる」と言った方が正確ではないだろうか。
・プランター内の生体は土壌微生物だけであり、光子エネルギーを利用できるのは生体は光合成細菌しかない。
消去法ではあるが、放射線の光子エネルギーを吸収しているのは光合成細菌である、と断言したい。
光合成細菌が、その密度を高め、活動が活性化するほど、放射線の吸収量は増加する、ということである。
しかし、この「光合成細菌の放射線吸収説」を裏付けるには、難儀するねぇ・・・、
ネットのどこを探しても「光合成細菌はγ放射線を吸収する」という情報は見当たらないのだ
3. 光合成細菌による光合成のメカニズム
そこで、光合成細菌がγ線を吸収できるという根拠を探るため、光合成のメカニズムをまとめてみた。
結論を先に言うなら、
・光合成細菌の基質(=エサ)は、光(光子エネルギー)と二酸化炭素である、これを原料として糖を合成する。
・γ放射線は、可視光線よりも遥かに強い光子エネルギーもっている。
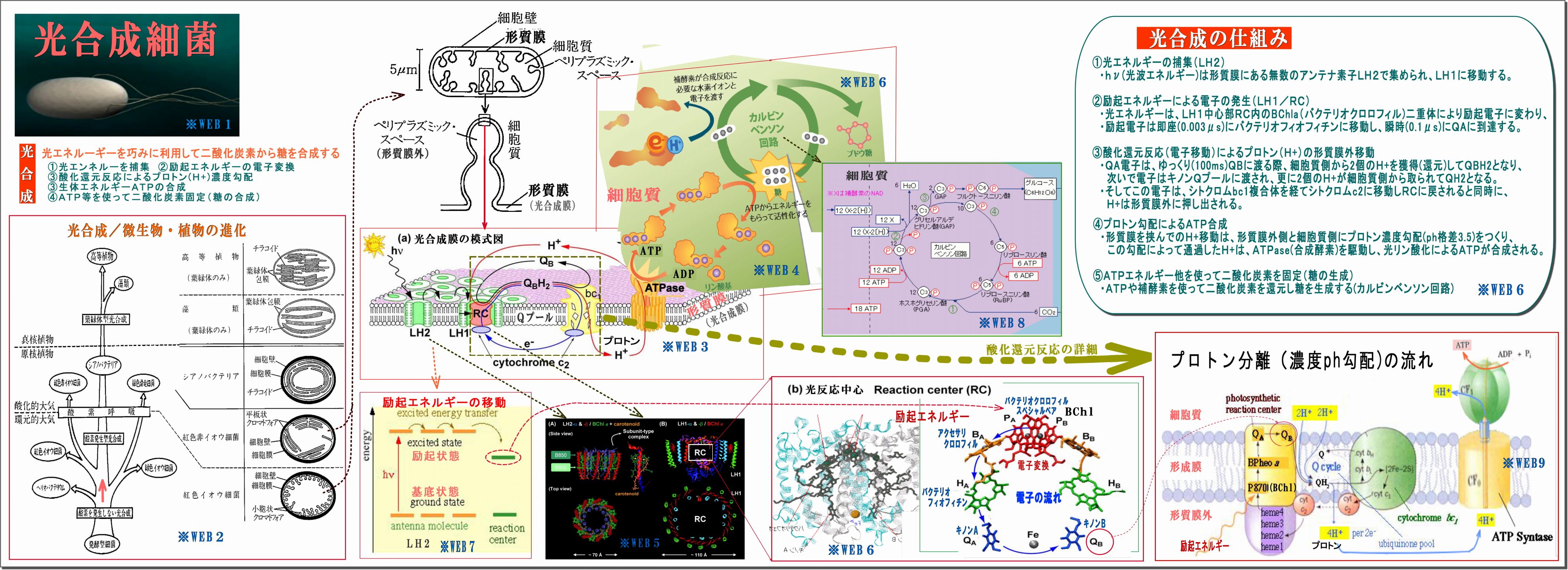
*WEB1 *WEB2 *WEB3 *WEB4 *WEB5 *WEB6 *WEB7 *WEB8 *WEB9
(1) 光合成細菌の細胞膜(形質膜:光合成膜とも言う)には光合成機能が詰まっている
・細胞膜が細胞内に向って小胞上のくびれ(クロマトフォア)がたくさんある。
この図では紅色イオウ細菌のものだが、紅色非イオウ細菌の場合は、クロマトフォアが
細胞質内に更にツリー状に広がっている。
・そのクロマトフォアには、たくさんの「光合成機能セット」が詰まっている、図はその一セットを拡大したもので、
特に光合成膜の面積大半を占めるのは、光エネルギー取り込むアンテナ素子LH2である。
(2) 光合成とは、光エネルギーを巧みに利用して二酸化炭素から糖を合成すること。 その流れは、
① 光エネルギーを捕集
・光で励起された光エネルギーをできるだけ広く集めるためのアンテナ素子がLH2であり、
・その励起エネルギーはLH1の反応中心RC(Reaction Center)に向って伝達される。
・励起エネルギーの伝搬方法は、励起とは軌道電子がより高い軌道に移動することであるが、
その電子は、瞬時に元の軌道に戻る際に他の原子の軌道電子を励起するという。
・この励起エネルギーはLH2→LH1→RCに瞬時に伝搬するらしい。
② 励起エネルギーの電子変換
・励起エネルギーはRCの中核BChl(バクテリオクロロフィル)にて電子に変換され、
バクテリオフィオフィチン→キノンQAへと瞬時に伝達される
③ 酸化還元反応によるプロトン(H+)濃度勾配
・QA電子はQBおよびキノンQプールに移動する際、細胞質側からプロトンH+を取ってQH2となり、
・電子は、後でシトクロム複合体を経てシトクロムc2からRCのヘムに戻されるが、
その前に、H+は形質膜外に押し出され、形質膜の内外にプロトン濃度(ph)勾配ができる。
④ 生体エネルギーATPの合成
・このプロトン濃度(ph)勾配によって、H+はATPase(合成酵素)を通過することで駆動され、
細胞質側で光リン酸化によってATPが合成される。
⑤ ATPを使って二酸化炭素固定(糖の合成)
・細胞質内では、ATPや補酵素を使って二酸化炭素を還元し、糖を合成する(カルビンベンソン回路)
3.光合成細菌が吸収できるのは可視光線だけなのか?
そんなことはない。
・いまや光合成細菌の吸収スペクトルは、研究され公表されているが、
この吸収スペクトルは、可視光線を照射して特定した「光学的スペクトル」のことであり、
光子エネルギー全般について特定されたものではない。
・光合成細菌は、光子エネルギーを吸収するメカニズムが備わっており、
γ放射線やX線などの光子エネルギーをも利用することができる。
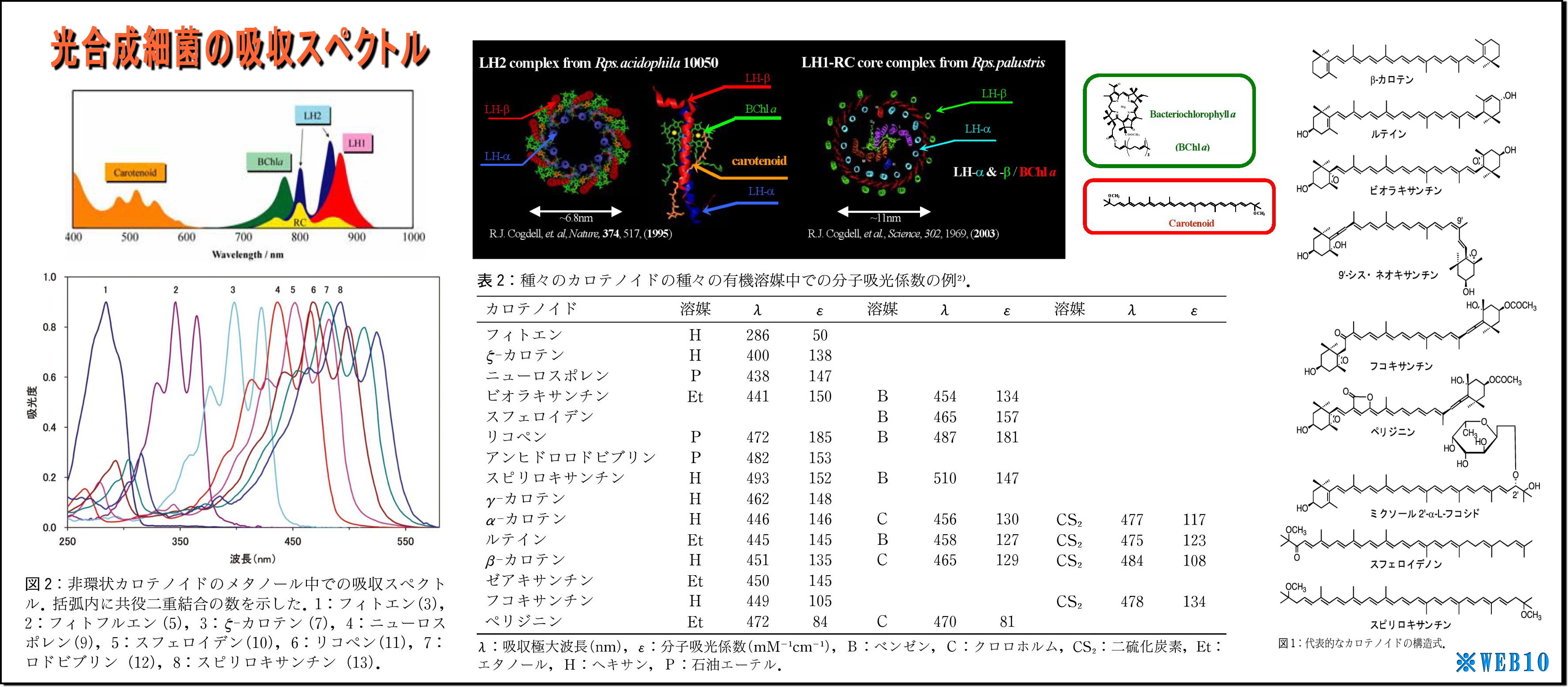
*WEB10
(1) LH2の立体構造を見ると、ヘリカル(螺旋)構造が目についてくる
・これは光も電磁波なので、光を効率よく集めるためなのだろうか、実に見事な形状である、
・また、このLH2の直径は70Å(オングストローム、1Å=0.1nm)=7nm程度であり、ヘリカル径はもっと小さいことから、
可視光線波長(390~750nm)向けアンテナ形状にしては小さいような気もする。
・そもそも原子の大きさは数nmなので、波長と共振できる形状だけから言及するなら、
分子レベルの大きさと構造(すなわち原子配列・形状・光入射向き)や色素複合体などとの関係が重要なのだろう。
・図の「吸収スペクトル」をみると、
・分子構造が複雑で大きいほど赤外線などの長波長に対応しているようであり、
・集光エネルギー終着点であるBChl(バクテリオクロロフィル)もスペクトルを持つが、
これはもっぱら光励起エネルギーの電子変換が主な役割らしい。
・またこれに絡み付くように繋がっているカロチノイドは短波長に対応するという、
・カロチノイドの基幹である共益二重結合の数Nが少ないほど短波長を吸収するようで、
N=3のフィトンでも280nm付近(UV-B中紫外線)止まりのようだ。
以上のことから、
吸収(エネルギー利用)できる光の範囲は、近紫外線~近赤外線(波長300~1,000nm)であるとされており、、
波長0.00187nmであるCs137のγ放射線は、吸収スペクトルから大きく外れていることになる。
このことをもって、「光合成細菌は放射線を吸収できない」「利用されない」と断言する人もいるようだ、
が、果たして、そう断言できるのか?
(2) しかし、私の実験では「γ放射線は光合成細菌によって吸収されている」という実証データがある。
① 公表されたスペクトル論に異を唱えるつもりは毛頭ない。
しかし、もっと短い波長の光子は吸収できない、とは書かれていない。
(3) 研究として公表されている「吸収スペクトル」の特定方法を見ると、
① タンパク質を溶媒に抽出し、近赤外線~可視光線~近紫外線の光を当てて調べたものである。
・つまり、単なる「光学的特性」の結果でしかない、ということだ。
・光は電磁波でもあるので、吸収スペクトルとは、本来電子回路系をも含めた周波数特性のことである。
電子回路でいうLCR(Lコイル、Cコンデンサ、R抵抗)のインピーダンスと周波数との共振性であり、
アンテナとは密接に関係するけど、決してアンテナ素子の物質・形状だけで決まるものではない。
・しかも、電子回路としてみた場合、一連の光化学プロセスは実に厄介な構造であり、いまだ解明されない処も
随所にあることから、周波数スペクトルを特定するには、実際に電子を流して検証する必要がある。
しかし、それは一切なされていない。
②更には、実際にX線や放射線を当てて吸収スペクトルを調べた訳ではもちろんない。
・地球に降り注ぐ太陽エネルギーの大半は可視光線とその前後だから対象外なのだろう。
(参考までに)いまの太陽光線中の割合は、およそ、
遠紫外線1%以下、近紫外線8%程度、可視光線40%、近赤外線20%、中赤外線25%、遠赤外線6% らしい。
・それとも、放射線の強い透過力では、吸収の変化は計測不能に近いからなのだろうか・・・。
・なお、LHCのヘリカル構造の直径は数Åであり、何やらX線帯近くまで対応できているような気がするが。
③そもそも、アンテナ素子LHC(Lh2、Lh1)は、光子エネルギーを受けて電子励起するものである。
・光子エネルギーは何も可視光に限らない、遠紫外線~X線~放射線のエネルギーは、
可視光の数百倍~数万倍~従十万倍もの強い光子エネルギーを持っているものなので、
放射線がLHCを通過する(軌道電子と衝突する確率は低い)だけで容易に軌道電子は励起されよう。
・つまり、アンテナ素子の元素配列や形状(光学的スペクトル)とは無関係に励起される。
・太古の地球では地表に降り注ぐ太陽エネルギーは陽子線・中性子線、X線や放射線が多かった事からして、
その時代に出現した光合成細菌は、生いたち初期からエネルギーの強い放射線やX線を利用していたと思われる。
LHCのヘリカル構造や分子の元素構造や配列形状は、放射線やXが少なくなる時期と合わせて進化したのだろう、
実に巧みに設計されたものと思われる。
④ 放射線の照射で細菌は死滅するということへの反論
・エネルギーMeV以上の強い放射線を局所集中的に照射するなら、死滅すると思われる。
・LHC構造のカロチノイドには、強い照射に対する防御機能(活性酸素を打ち消す役割)もあるらしい
・私の実験では、光合成細菌や酵母などの共生菌が死滅または増殖が阻害されているとは考えられない、
むしろ元気なくらいに感じられる。
・チェルノブイリ事故周辺やフクシマ現場周辺の写真を見る限り、植物が赤茶けて枯れた様子は全く見られない。
・光合成の備わった生物は、放射能に強く、光合成も盛んになるもの、と思われる。
⑤ なお、X線や放射線の吸収性(スペクトル)を確かめたいならば、
・細菌が死んだ状態では計れない、単に透過するだけだから、生きてシステムが働いている状態で測定しなければならない。
・なお、例え細菌が生きていたとしても、負荷が必要である、負荷とは出力負荷=消費のことだ、
つまり、合成されたATPを消費してあげないと、飽和して励起エネルギーの流れはすぐに停滞してしまう、
・電気実験だって、いくら電圧をかけても電力が消費されなければ電流は流れない。
・ATPが消費され、合成された糖などが共生菌によって消費され、更に植物草に消費される、など、
一連の微生物叢や植物との共生が出来上がって、はじめて放射線の吸収が確認できるものと考えたい。
この観点から、私の実験では、詳細なスペクトルまでは調べることはできないが、
間違いなく「放射線を吸収している」ことがを実証しているのである。
4. 光合成細菌の放射線吸収の謎にせまる
光合成細菌は、太古の地球が太陽風(陽子線・放射線・紫外線に晒されていた環境で出現し、進化してきた微生物なので、
放射線に強いというよりも「放射線を利用できるメカニズム」が備わっているものと推測する。
① 光合成細菌に光エネルギー捕集と電子変換の見事なメカニズム
・実に見事に設計されているものと感嘆する。
(イ) 電磁波エネルギーと光子エネルギーの両方を利用できるメカニズムとなっている
(ロ) 電磁波エネルギーは、広く配置したアンテナ素子LHCで捕集する機能、
可能な限り広範囲な可視光線帯のエネルギーを吸収しやすいために
クロロフィルや多様な分子構造をもつカロチノイドの構造と配置によるスペクトルの形成
(ハ) 可視光線より遥かに波長の短い光子(遠紫外線・X線・γ線)のエネルギーは、
光学的スペクトルとは無関係に、直接LHCを励起する。
(ニ) 可視光線近辺の光捕集による励起エネルギーも、より短波長の光子による励起エネルギーも、
バクテリオクロロフィルPにて同一視して電子変換されてる
(ホ) 電子のキノンと結びつき、ATP合成のためのプロトン勾配を作る
(ヘ) 生体エネルギーATPを利用して糖を合成する
・実に見事としか言いようがない。
② 地球の進化は生物の進化と共にある
・地球という惑星が誕生したのは46億年前で、最初の生命体誕生は、今なお謎らしい。
38億年前の岩石には、すでに生命の痕跡が見つかっている。
・最初の生物は微生物だが、他から栄養をもらって増殖する従属栄養微生物が多いなか、
自ら栄養を作り出せる独立栄養微生物は光合成細菌だけである、
・この光合成細菌の誕生(微生物の進化)によって、地球は飛躍的に生命力が増してきた。
・光合成細菌の最初は、酸素を発生しない光合成細菌であった。
有毒ガスである硫化水素の多い地球だから、H2Sを還元して光化学する紅色(や緑色)硫黄細菌であり、
次第にH2Oを還元し酸素を排出する非硫黄細菌へと進化する。
・この光合成細菌の光合成機能を受け継いだ原核植物シアノバクテリア(藍藻)へと進化し、
ついには植物の葉のなかの葉緑体へと発展していく。
・この進化のようすはクロマトフォアの密集度合いの推移にみることができる。
・なお余談だが、光合成の中間産物である生体エネルギーATP機能は、動物細胞のミトコンドリア
として細胞内に組込まれていく。
③ 太古の地球には酸素はほとんど無く、やがて活発なマグマが落ち着くころになると、
・地殻が安定し、自転による地磁気が明確になってバンアレン帯が形成される27億年前ころになると、
陽子線や放射線などが含まれる有害な地球への太陽風は次第に弱まり、
やがて酸素の層が厚みを帯びてきて、10億年前頃にはオゾン層ができると紫外線も弱まり、
太陽エネルギーは可視光線がメインとなってきた。
④ 光合成細菌は、このバンアレン帯が出来上がる以前に登場し進化を続けてきた
・バンアレン帯以前の地球は、陽子線や放射線など生体に有害エネルギーが降り注ぐ状態である、
こんな環境でも、光合成細菌のタンパク質などは破壊されないで進化・増殖できたのには、
放射線をも基質(増殖に必要なエサ)として利用してきたと想像する。
・可視光線の数十万倍ものエネルギーのある放射線を、生命の増殖に使わない手はないし、
過酷な環境に順応し、逆に過酷な環境を好むように光合成細菌は進化してきたものと素直に考えたい。
⑤ 光合成細菌の進化について
・上記①に述べたように、光合成の見事なまでの光化学メカニズムは、いきなり完成したわけではない。
・複雑な光学的スペクトルが形成できたのは、進化のずっと後のことで、放射線よりも可視光線の割合が
多くなってきたことに適応しながらの進化ではなかろうか。
・つまり、地球太古の時代は、可視光線よりも遥かにエネルギー高いX線やγ線が降り注いでいたので、
簡単に励起エネルギーを得ることができるシンプルな光化学系メカニズムが最初にできあがった、と考えたい。
そして、バンアレン帯以降あたりから可視光線をも吸収しやすいような光学スペクトル分子構造へと進化してきた。
以上、私がいろいろ調べてみたことを、私の推測も交えて、備忘録的に取りまとめてはみた。
だいぶ謎の核心に近づいてきたのではないかと思っているが、如何なものだろうか