
放射線吸収説とその実証
投稿ツリー
-
 放射能低減の実験報告 (放射線吸収・放射能減量を実証) (go, 2014/10/31 18:59)
放射能低減の実験報告 (放射線吸収・放射能減量を実証) (go, 2014/10/31 18:59)
-
実験方法およびデータ処理 (go, 2014/11/1 6:41)
-
放射線吸収説とその実証 (go, 2014/11/1 6:42)
-
放射能崩壊加速説?現象の確認 (go, 2014/11/3 3:55)
-
実験を終えて (go, 2014/11/18 10:54)
-

放射線吸収説とその実証
msg# 1.2
go
 投稿数: 125
投稿数: 125
投稿数: 125
光合成細菌と放射能の関係について、専門家たちの見解は、
「光合成細菌は、放射線は吸収できない」
「利用できるのは可視光線前後(近赤外線~近紫外線)だけである」
というものだ。
これが巷のEMパッシング(とんでも科学)の背景の一つなっていると思われる。
私は、2013年夏に「実験そのⅠ」を行い、「光合成細菌は放射線を吸収する」という現象を捉えて公開した。
翌2014年夏には、再現実験「実験そのⅡ」を行った結果、この「放射線吸収説」の正しさの確証を得ることができたので、
以下にまとめてみた。
1。光合成細菌による放射線吸収メカニズムについて -------------
(1) 吸収スペクトルについて
植物の葉緑体や光合成細菌は太陽光エネルギーを利用して光合成を行うが、多くの研究の結果から導かれた
吸収スペクトルを根拠に「光合成細菌は放射線を利用できない」と言う人たちがいるが、これは正しいのか?
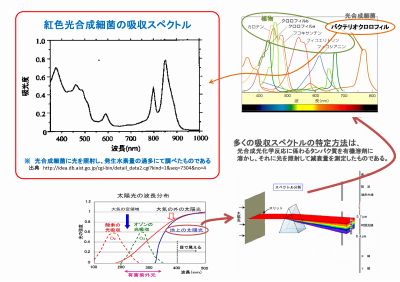
吸収スペクトルを調べてみると、
測定する光源の波長は地上降り注ぐ
太陽光のスペクトルの範囲だけであった。
右図左上は、紅色光合成細菌の吸収スペ
クトルだが、活きた光合成細菌に光を
当てて発生する水素の量を調べたもの
としては非常に貴重なデータである。
しかし一般的な吸収スペクトル(図右上)
は、溶剤に溶かした光化学反応に係わる
タンパク質の透過量を測って求めたもの
なので、分子構造と元素配列に依存した
「光学的なスペクトル」でしかない。
このような方法による吸収スペクトを根拠にして、太陽光より遥かに波長の短いX線やガンマ放射線は
「光合成細菌は光合成に利用でりはずはない」と断定することは、大いに間違いである。
光合成細菌は放射線を吸収し利用できることを確かめるのは難しいように思われる。
・太陽光より遥かに波長の短いX線や更に短いガンマ放射線を死んだ光化学反応タンパク質に照射しても
透過力が強いだけに、単に透過するだけだと思われるし、、
・光化学反応の微弱な電子の流れを測定すればいいことだが、これはなお難しいように思われる。
しかし、私の行った実験そのⅠ・そのⅡの方法によれば、
光合成細菌が放射線をが吸収している、という現象が確認できるのである。
(2) 光合成細菌の光反応メカニズムについて
はたして、光合成細菌には放射線を吸収できるメカニズムが備わっているのだろうか?
すでに、光合成細菌の光合成(光化学反応および糖合成反応)のメカニズムは詳しく解明されている(下図)。
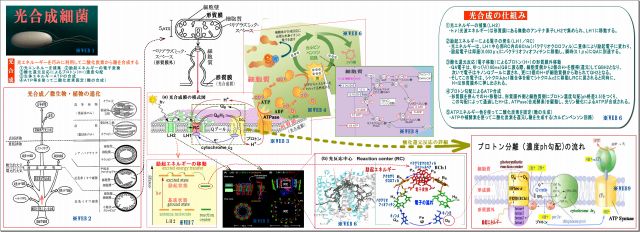
光合成全体のメカニズムは、
・アンテナ素子LH2が光子エネルギーによって励起される
・励起エネルギーはRCに向って伝搬しながら移動し、RCバクテリオクロロフィル(スペシャルペア)
によって電子化される。
・この電子はキノンQに伝わり、プロトンH+が光合成膜の外側に生まれる。
・光合成膜の内外のプロトン勾配によってATPが合成され、
・二酸化炭素を固定して糖合成へと進む
ここで着目したいことは、
① 利用する光は、広く「光子エネルギー」であって、「太陽光エネルギー」に限らないということである。
つまり「励起できる強さの光子エネルギー」があればいいということだ。
② 光子エネルギーの強さは「波長の逆数に比例する」、つまり波長が短いほど光子エネルギー大きい、
セシウム光子波長は太陽光の30万分の1なので、セシウムは太陽光の30万倍もエネルギーが強い、
つまり、X線でも放射線なら太陽光より容易に励起されうることになる。(参考)「励起についての補足」
ということで、光合成細菌は「放射線を吸収できない」とするのは全くの誤りである、
「放射線を吸収し利用できる」と言うことである。
現実では、私の行った実験そのⅠ・そのⅡ法によって、放射線が吸収されているという現象を確認している。
(参考)「励起についての補足」
太陽光も含め、それより波長の短い電磁波は光子ともよばれ、本来は電磁波なのだが粒子のような性格も併せ持っている。
(a) 粒子としてのエネルギー励起について
・X線や放射線は物質と相互作用(コンプトン効果、光電効果)を引き起こす。
放射線が物質に当たると、軌道電子を起動外に弾き飛ばしだけでなく低いエネルギーの放射線が
散乱する(コンプトン散乱)。
・この散乱放射線は、次々とコンプトン効果を繰り返すが、あるレベルまでエネルギーが低下すると
放射線は発せず、単に電子だけが飛び出すか、軌道電子が励起される(光電効果)
・このことにより、植物葉緑体のクロロフィルや光合成細菌のバクテリオクロロフィルはエネルギー励起される。
(b) 電磁波としてのエネルギー励起について
・光合成細菌の受光素子LH2/LH1の立体構造と電子発生の流れから電子回路であり、
ヘリカル(螺旋状)なポリペプチドや円形構造は電子部品のコイルを彷彿させる、
まるで精巧な電磁波の受信回路と考えることもできる。
・LH2などの寸法は、概ね10Å(オングストローム、1Å=0.1nm)界隈なので、電磁波波長と比較するなら、
太陽光の500分の1程度であり、X線領域である。
・電磁波のアンテナ素子とするなら、この寸法の数分の1~数百分の1の電磁波波長を捉える能力があり、
X線やγ放射線は電磁波でもあるので、共鳴現象は可能である。
・もし回路側のインピーダンスが入力波長とマッチングして電磁共鳴できるのなら、
もっとも効率的にエネルギーを得ることができる。
・光合成細菌は生き物である、環境の変化に応じてアンテナ素子のインピーダンスを微調整することは
可能であろう。微調整と言ってもタンパク質の構成を変える必要はなく、ほんのわずかメタボまたはスリムに
なるだけでインピーダンスは変われるからだ。
例えば、セシウムからのγ線が多い環境なら、その波長に合わせて微調整するか増殖するだろうから、
放射線の電磁波エネルギーを効率よく取り込み、励起エネルギーを得ることができよう。
光合成細菌は、放射線エネルギー能動的に取りこむことができる原始的な生き物であると想像する。
2.光合成細菌による放射線吸収説を裏付ける現象 ---------------
下図は、「実験そのⅠ」および「実験そのⅡ」のものである。 両実験とも、
・実験前半は光合成細菌が増殖し活性化すると思われるさまざまな促進資材の投入を試し、
・実験の後半では土壌を自然乾燥させて微生物の活性を弱めるなどして観察し、
・最後は、土壌全体を加熱乾燥して、微生物に影響されない土壌本来の放射能による放射線量を調べた。
・実験そのⅡでは、土壌の加熱乾燥処理は、実験開始時と実験終了時におこなった。
・また、実験そのⅡでは、実験終了時の土壌を加熱乾燥する直前に、土壌全体を撹拌して放射線量の変化を調べた。
以上の実験の結果、光合成細菌は放射線を吸収している現象を確認したので、以下に説明する。
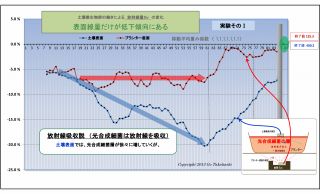
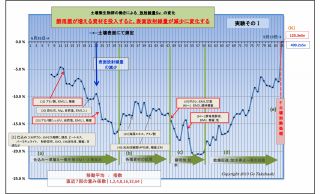
(1) 土壌表面および底面の放射線量の変化の違い (放射線吸収説の裏付け-1)
実験そのⅠおよびそのⅡでは、共に、
・底面の変化(赤線)は概ね横ばいであるのに対し、
・表面の変化(青線)は微生物が働くにつれて下降(放射線量が低下)している。
この現象は、実験の経過と共に、土壌表面に光合成細菌が増殖したことによって、
土壌から発する放射線は光合成細菌層に吸収されて低下したものと考えられる。
ここで一呼吸して、土壌表面線量が低下する他の一般的な要因についてガン変えてみたい。
① 土壌に含まれる水分の過多によても線量は変化するものである。
実験開始数日後からは、土壌水分が飽和状態になることを意識してし加水している。
(プランター受け皿に水が少し染み出るほどにして、溢れた水は翌日にプランターに戻す)
このように土壌水分はほぼ一定に保ってはいるはずだが、土壌表面放射線量の低下/上昇の変化のようすから、
水分量の過多だけによる変化ではないと言える。
② 土壌表面の測定器の位置が土壌面から離れることによって線量が弱まるものである。
確かに少しづつは離れていき、それによって線量が低下することは確かである。
しかし土壌表面の変化のようすをよく見ると、実験そのⅠでは大きく上昇している箇所もある。
実験そのⅡでは、ボカシなどの投入量に必ずしも比例しない放射線量の変化も随所に見られることから、
距離以外の要因もあるものと考える。
③ 土壌中のセシウムが徐々に底面方向に移動しているのではないか?
それならば、底面放射線が徐々に上昇して現象には合わないし、自然乾燥時の変化を見る限り、
セシウムの沈下は有意には表れてはいない。
以上のことから、土壌表面の表面線量の低下や変化は、土壌微生物の働きが深く関与しているものと考えられる。
なお、多様な微生物群のうち、光子エネルギーを吸収利用できる微生物は「光合成細菌」しかないことから、
放射線の土壌表面での低減は光合成細菌によるものに違いない考える(放射線吸収説)。
(2) 光合成細菌は多量に増殖している (放射線吸収説の裏付け-2)
 この写真は、実験そのⅡのプランター内壁面のものだが、
この写真は、実験そのⅡのプランター内壁面のものだが、
光合成細菌(赤色)がびっしりと増殖し付着している。
壁面内側には太陽光の直接の照射はないので、
壁面の熱によって増殖がしたものと思われが、
底面にはそれもなく光合成細菌は全く増殖していない。
このような状況から、光合成細菌はプランター内でも増殖していることが確認できる。
土壌表面の光合成細菌は迂闊にも確認しなかったが、太陽光や反射光が最も多く直接照射する所なので、
内部壁面よりも遥かに多量の光合成細菌は層を形成していると考えられる。
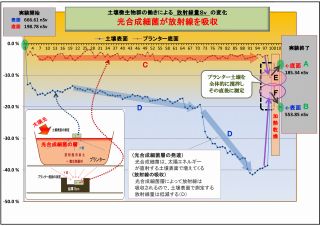
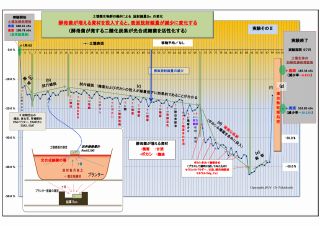
(3) 放射線吸収説を裏付ける土壌撹拌後の放射線量の変化 (放射線吸収説の裏付け-3)
「実験そのⅡ」において、実験終盤で「プランターの土壌全体を撹拌した直後に測定」したところ、
放射線量が大巾に変化しているが、これは次のように説明できる。
① 土壌を撹拌したことで、表層に集中していた光合成細菌が土壌全体に散らばったので、
底面方向に移動した光合成細菌によって放射線は吸収され、結果として放射線量が低く測定された(E)
② 同じ理由により、表面に集中していた光合成細菌が大幅に減ったので、土壌放射線の吸収量が激減し、
表面の放射線量は大幅に上昇した(F)
この二つの現象は、光合成細菌による「放射線吸収説」を強く裏付けるものであり、物的証拠となるデータでもある。
(4) 二酸化炭素を供給すると光合成細菌が活性化する (放射線吸収説の裏付け-4)
実験そのⅠおよび実験そのⅡの土壌表面に変化を再度注視していただきたい。
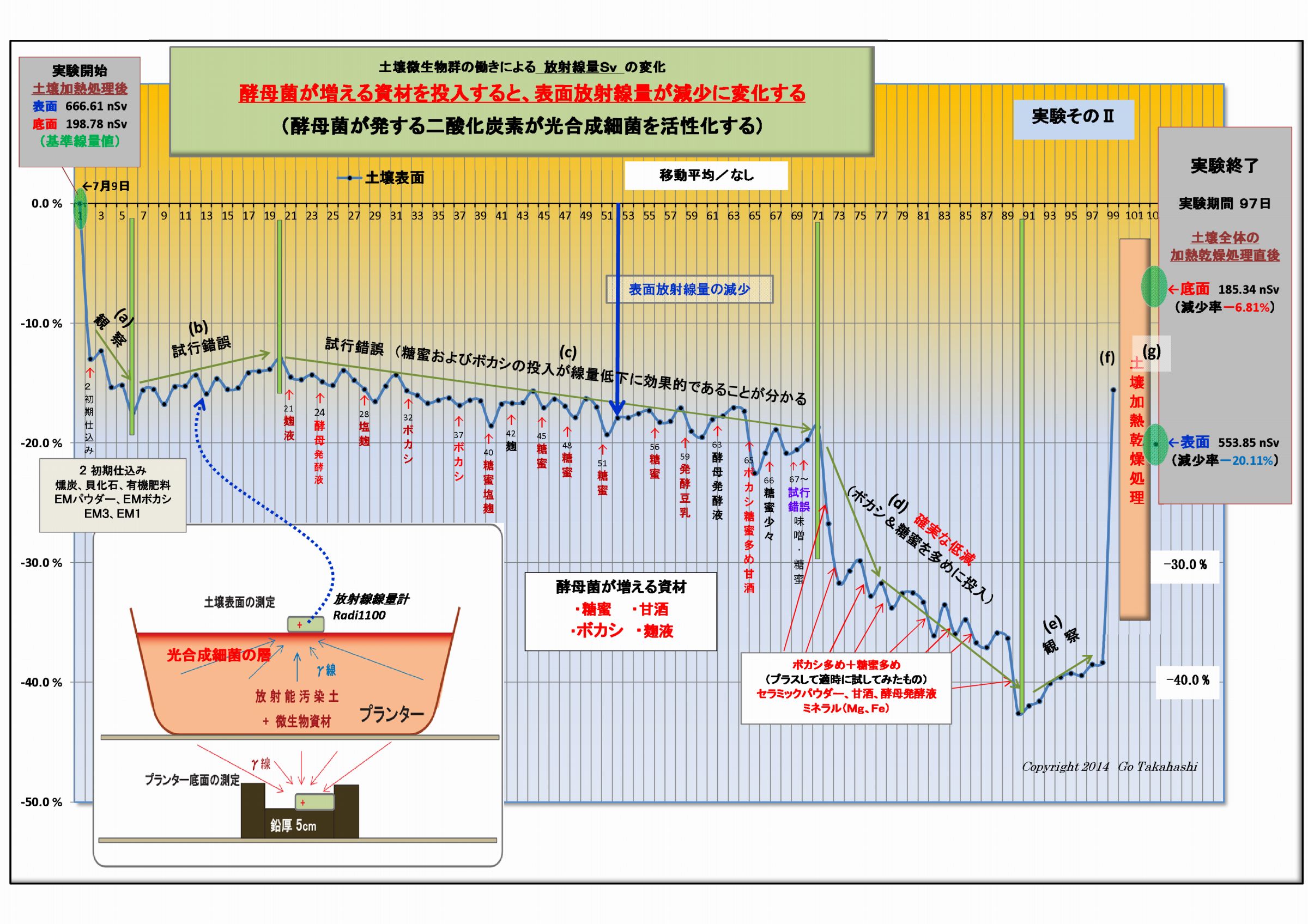
酵母菌が増えるような資材を投入すると、土壌表面放射線が減少する、という因果関係な現象が読み取れる。
(実験では、土壌表面の放射線量を低下させるために意識的に酵母菌の活動を強化した)
米ぬか発酵ボカシや麹に酵母菌が多く含まれており、糖蜜や甘酒(麹由来の)は酵母菌が増殖するための
優良なエサであり、甘さの残る酵母発酵液は酵母菌が多量に増殖している。
光合成細菌は酵母菌と相性がいい、酵母菌から発生する二酸化炭素は光合成細菌には「不可欠なエサ」であるからだ。
下図は、光合成細菌の生体内部の機能をまとめたものである。
光合成細菌は、二酸化炭素CO2と光子エネルギーをエサとして糖を産生する微生物である。
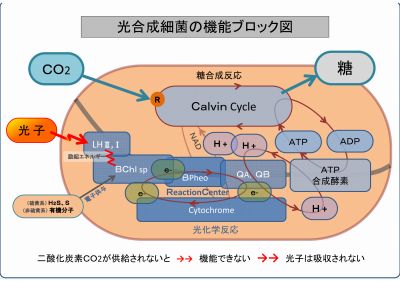
光合成細菌は、二酸化炭素CO2がなければ働けない・機能できない。
実験データによる土壌表面の放射線量の変化を注視すると、酵母関連資材を投入すると表面放射線量は低下し、
酵母菌が少ないと表面放射線は反転上昇するようすが見て取れる。
つまり、光合成細菌は、二酸化炭素CO2が供給されないと光子エネルギーを吸収できないのである。
このことを、ブロック図の機能を出口方向から順に説明する。
・CO2が無いと、糖を合成できないだけでなく、カルビン回路そのものが循環しない
↓↓↓↓
・ADPが還流されないので、ATP合成酵素は働けない
↓↓↓↓
・プロトンH+の循環が止まる(飽和状態)
↓↓↓↓
・e-電子の循環が止まる
↓↓↓↓
・シトクロームからの電子の還流が無いと酸化したバクテリオクロロフィルの機能は復元できない
↓↓↓↓
・光子からのエネルギーは吸収されない => 放射線は吸収できない
つまり、二酸化炭素の供給がなければ光合成細菌は光子(放射線)エネルギーを吸収できない、ということである。
( 身近な例だが、電力は柱上トランスの電磁誘導を経由して二次側の家庭に引き込まれているが、
電気が消費されなければ電流はながれないので、高圧線の電力は消費=吸収されない。
光合成細菌でいうなら、糖合成に必要なの原料が消費されないと光子エネルギーは吸収されない原理と同じである )
これが光合成細菌による「放射線吸収説」の正しさを説明できるメカニズムであり、
実験で投入した酵母促進資材の「不足/充足」と表面線量の「上昇/低下」の因果関係の説明にもなっている。
光合成細菌単独では働けない、光合成細菌を中核とした良好なマイクロバイオーム(微生物叢)が放射線の吸収・利用を高めている。
「光合成細菌は、放射線は吸収できない」
「利用できるのは可視光線前後(近赤外線~近紫外線)だけである」
というものだ。
これが巷のEMパッシング(とんでも科学)の背景の一つなっていると思われる。
私は、2013年夏に「実験そのⅠ」を行い、「光合成細菌は放射線を吸収する」という現象を捉えて公開した。
翌2014年夏には、再現実験「実験そのⅡ」を行った結果、この「放射線吸収説」の正しさの確証を得ることができたので、
以下にまとめてみた。
1。光合成細菌による放射線吸収メカニズムについて -------------
(1) 吸収スペクトルについて
植物の葉緑体や光合成細菌は太陽光エネルギーを利用して光合成を行うが、多くの研究の結果から導かれた
吸収スペクトルを根拠に「光合成細菌は放射線を利用できない」と言う人たちがいるが、これは正しいのか?
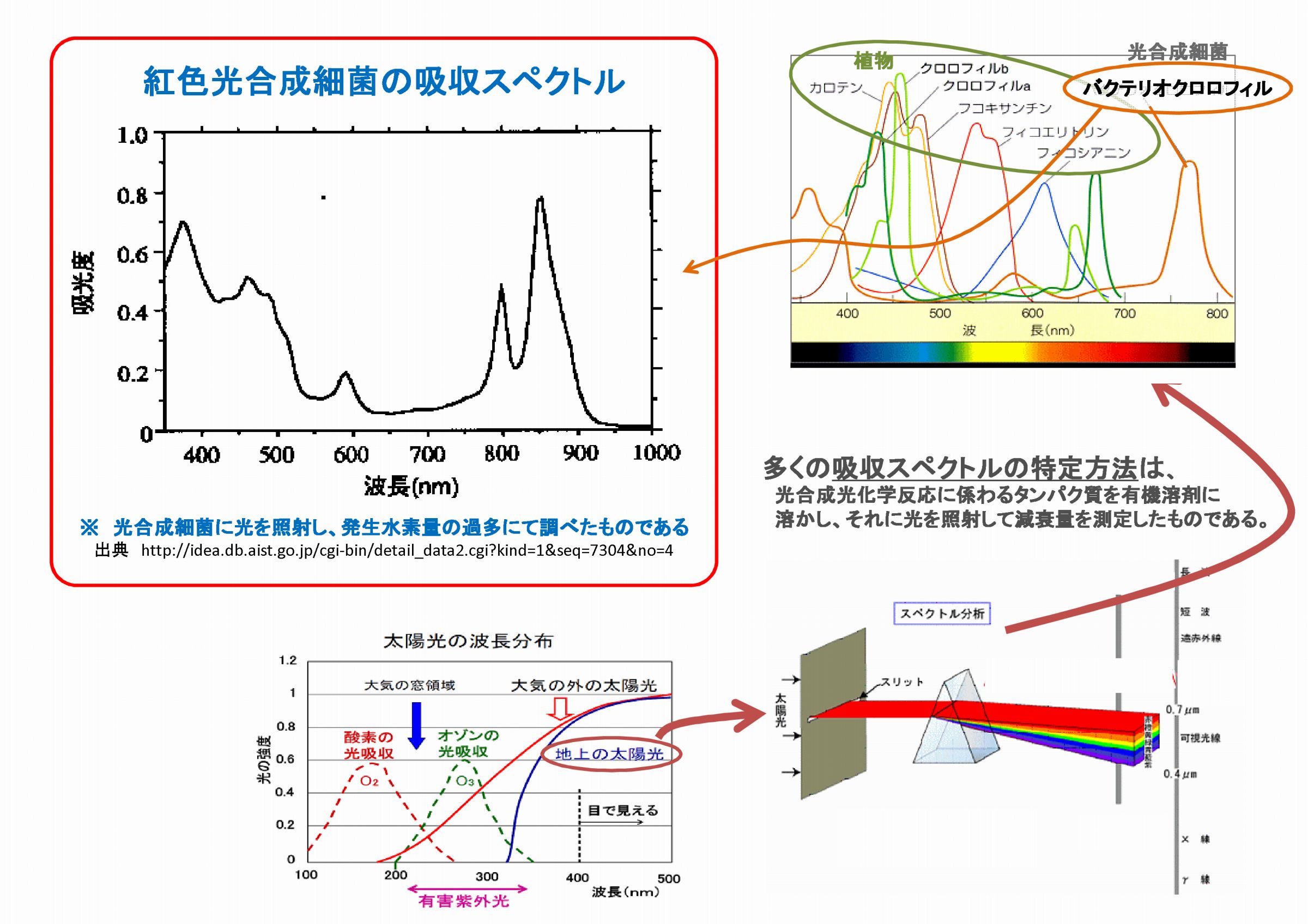
吸収スペクトルを調べてみると、
測定する光源の波長は地上降り注ぐ
太陽光のスペクトルの範囲だけであった。
右図左上は、紅色光合成細菌の吸収スペ
クトルだが、活きた光合成細菌に光を
当てて発生する水素の量を調べたもの
としては非常に貴重なデータである。
しかし一般的な吸収スペクトル(図右上)
は、溶剤に溶かした光化学反応に係わる
タンパク質の透過量を測って求めたもの
なので、分子構造と元素配列に依存した
「光学的なスペクトル」でしかない。
このような方法による吸収スペクトを根拠にして、太陽光より遥かに波長の短いX線やガンマ放射線は
「光合成細菌は光合成に利用でりはずはない」と断定することは、大いに間違いである。
光合成細菌は放射線を吸収し利用できることを確かめるのは難しいように思われる。
・太陽光より遥かに波長の短いX線や更に短いガンマ放射線を死んだ光化学反応タンパク質に照射しても
透過力が強いだけに、単に透過するだけだと思われるし、、
・光化学反応の微弱な電子の流れを測定すればいいことだが、これはなお難しいように思われる。
しかし、私の行った実験そのⅠ・そのⅡの方法によれば、
光合成細菌が放射線をが吸収している、という現象が確認できるのである。
(2) 光合成細菌の光反応メカニズムについて
はたして、光合成細菌には放射線を吸収できるメカニズムが備わっているのだろうか?
すでに、光合成細菌の光合成(光化学反応および糖合成反応)のメカニズムは詳しく解明されている(下図)。
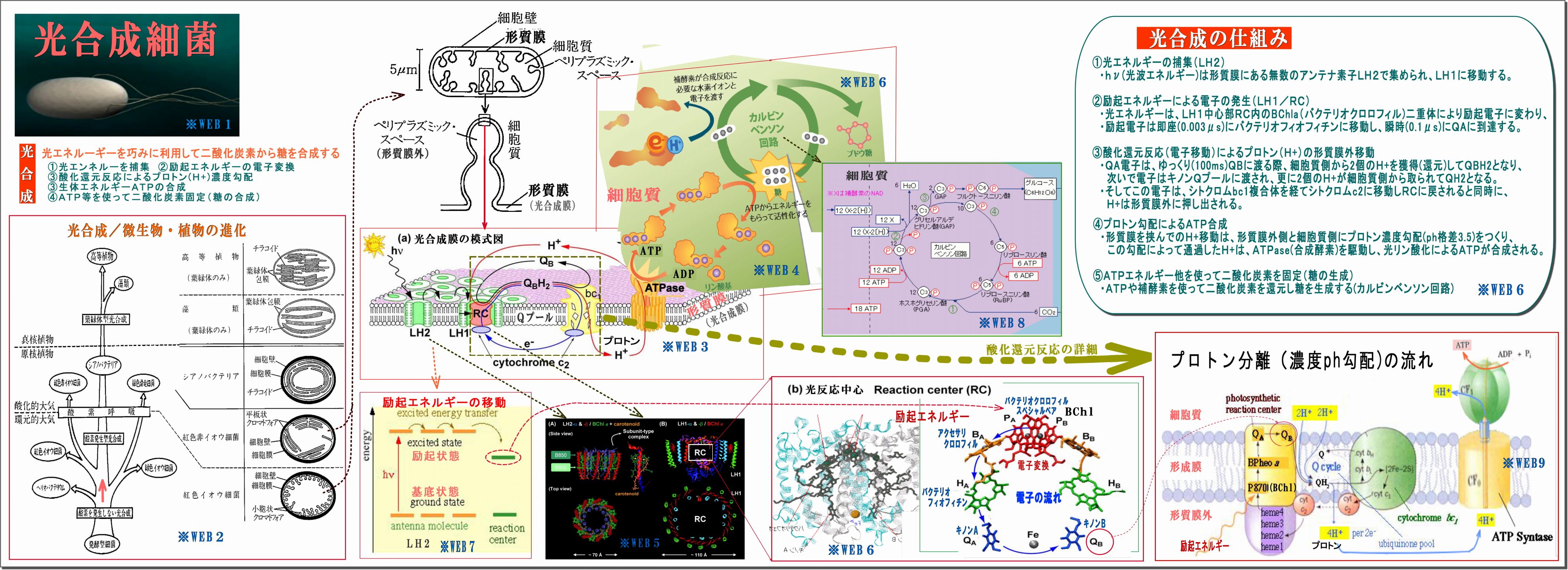
光合成全体のメカニズムは、
・アンテナ素子LH2が光子エネルギーによって励起される
・励起エネルギーはRCに向って伝搬しながら移動し、RCバクテリオクロロフィル(スペシャルペア)
によって電子化される。
・この電子はキノンQに伝わり、プロトンH+が光合成膜の外側に生まれる。
・光合成膜の内外のプロトン勾配によってATPが合成され、
・二酸化炭素を固定して糖合成へと進む
ここで着目したいことは、
① 利用する光は、広く「光子エネルギー」であって、「太陽光エネルギー」に限らないということである。
つまり「励起できる強さの光子エネルギー」があればいいということだ。
② 光子エネルギーの強さは「波長の逆数に比例する」、つまり波長が短いほど光子エネルギー大きい、
セシウム光子波長は太陽光の30万分の1なので、セシウムは太陽光の30万倍もエネルギーが強い、
つまり、X線でも放射線なら太陽光より容易に励起されうることになる。(参考)「励起についての補足」
ということで、光合成細菌は「放射線を吸収できない」とするのは全くの誤りである、
「放射線を吸収し利用できる」と言うことである。
現実では、私の行った実験そのⅠ・そのⅡ法によって、放射線が吸収されているという現象を確認している。
(参考)「励起についての補足」
太陽光も含め、それより波長の短い電磁波は光子ともよばれ、本来は電磁波なのだが粒子のような性格も併せ持っている。
(a) 粒子としてのエネルギー励起について
・X線や放射線は物質と相互作用(コンプトン効果、光電効果)を引き起こす。
放射線が物質に当たると、軌道電子を起動外に弾き飛ばしだけでなく低いエネルギーの放射線が
散乱する(コンプトン散乱)。
・この散乱放射線は、次々とコンプトン効果を繰り返すが、あるレベルまでエネルギーが低下すると
放射線は発せず、単に電子だけが飛び出すか、軌道電子が励起される(光電効果)
・このことにより、植物葉緑体のクロロフィルや光合成細菌のバクテリオクロロフィルはエネルギー励起される。
(b) 電磁波としてのエネルギー励起について
・光合成細菌の受光素子LH2/LH1の立体構造と電子発生の流れから電子回路であり、
ヘリカル(螺旋状)なポリペプチドや円形構造は電子部品のコイルを彷彿させる、
まるで精巧な電磁波の受信回路と考えることもできる。
・LH2などの寸法は、概ね10Å(オングストローム、1Å=0.1nm)界隈なので、電磁波波長と比較するなら、
太陽光の500分の1程度であり、X線領域である。
・電磁波のアンテナ素子とするなら、この寸法の数分の1~数百分の1の電磁波波長を捉える能力があり、
X線やγ放射線は電磁波でもあるので、共鳴現象は可能である。
・もし回路側のインピーダンスが入力波長とマッチングして電磁共鳴できるのなら、
もっとも効率的にエネルギーを得ることができる。
・光合成細菌は生き物である、環境の変化に応じてアンテナ素子のインピーダンスを微調整することは
可能であろう。微調整と言ってもタンパク質の構成を変える必要はなく、ほんのわずかメタボまたはスリムに
なるだけでインピーダンスは変われるからだ。
例えば、セシウムからのγ線が多い環境なら、その波長に合わせて微調整するか増殖するだろうから、
放射線の電磁波エネルギーを効率よく取り込み、励起エネルギーを得ることができよう。
光合成細菌は、放射線エネルギー能動的に取りこむことができる原始的な生き物であると想像する。
2.光合成細菌による放射線吸収説を裏付ける現象 ---------------
下図は、「実験そのⅠ」および「実験そのⅡ」のものである。 両実験とも、
・実験前半は光合成細菌が増殖し活性化すると思われるさまざまな促進資材の投入を試し、
・実験の後半では土壌を自然乾燥させて微生物の活性を弱めるなどして観察し、
・最後は、土壌全体を加熱乾燥して、微生物に影響されない土壌本来の放射能による放射線量を調べた。
・実験そのⅡでは、土壌の加熱乾燥処理は、実験開始時と実験終了時におこなった。
・また、実験そのⅡでは、実験終了時の土壌を加熱乾燥する直前に、土壌全体を撹拌して放射線量の変化を調べた。
以上の実験の結果、光合成細菌は放射線を吸収している現象を確認したので、以下に説明する。
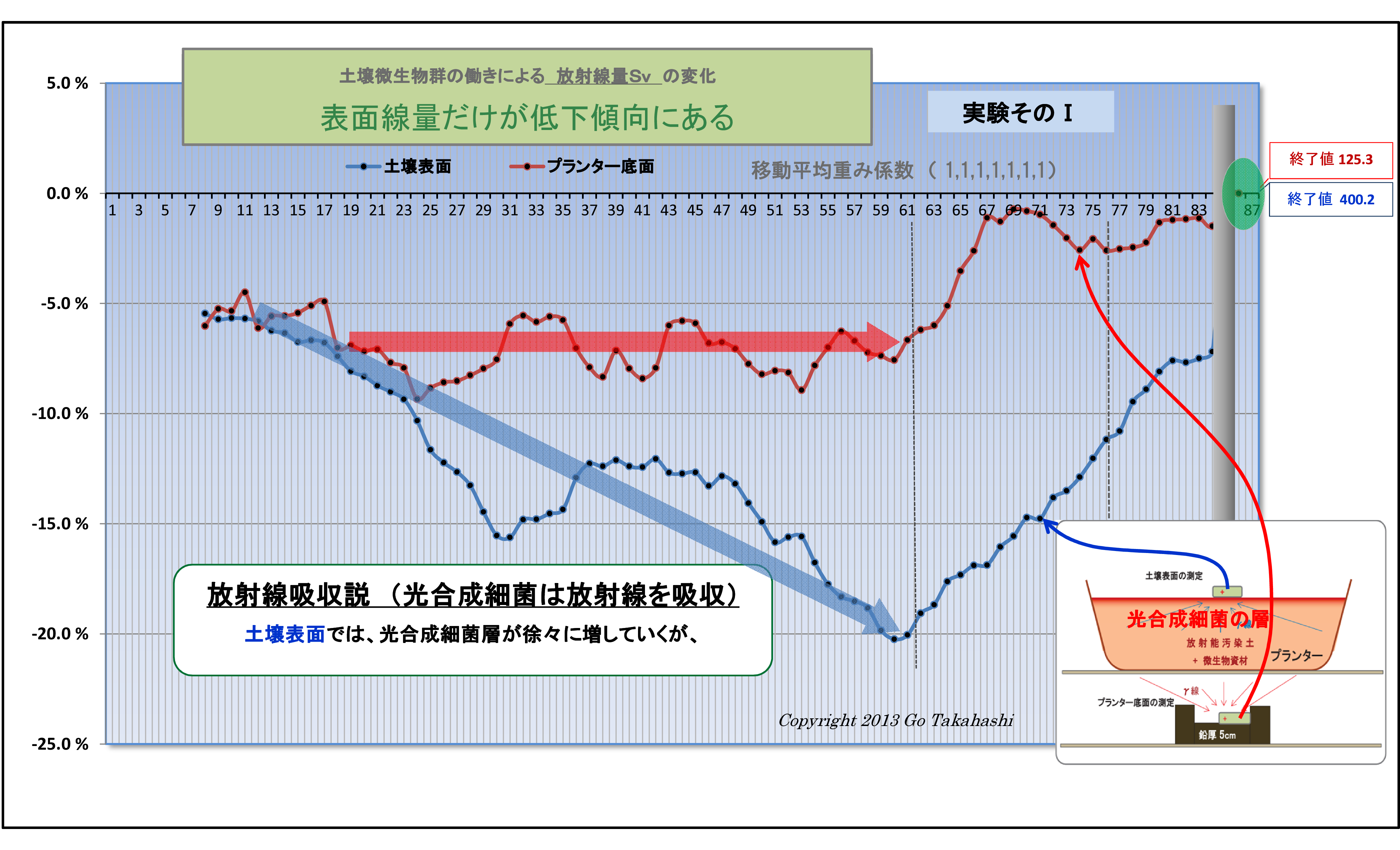
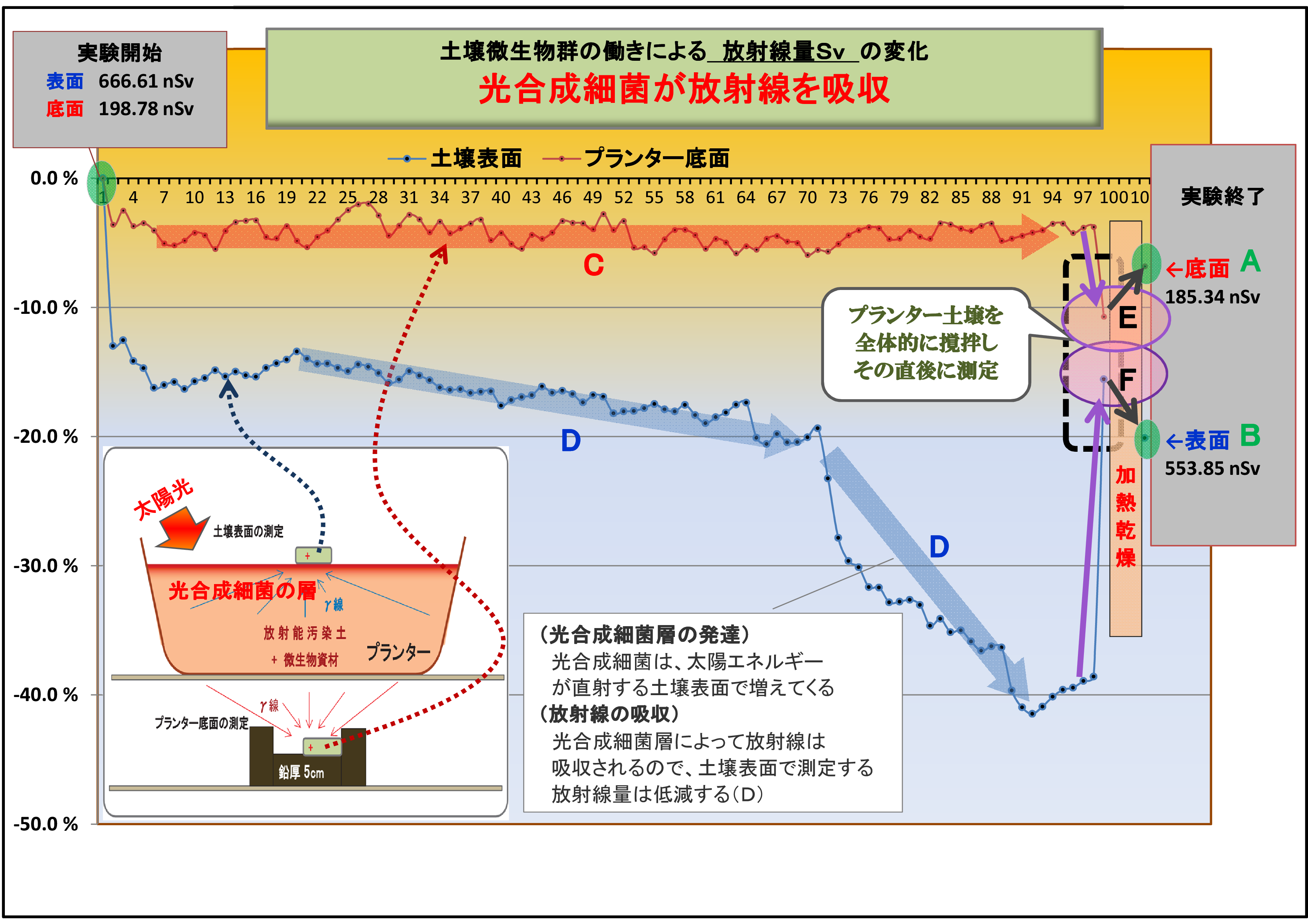
(1) 土壌表面および底面の放射線量の変化の違い (放射線吸収説の裏付け-1)
実験そのⅠおよびそのⅡでは、共に、
・底面の変化(赤線)は概ね横ばいであるのに対し、
・表面の変化(青線)は微生物が働くにつれて下降(放射線量が低下)している。
この現象は、実験の経過と共に、土壌表面に光合成細菌が増殖したことによって、
土壌から発する放射線は光合成細菌層に吸収されて低下したものと考えられる。
ここで一呼吸して、土壌表面線量が低下する他の一般的な要因についてガン変えてみたい。
① 土壌に含まれる水分の過多によても線量は変化するものである。
実験開始数日後からは、土壌水分が飽和状態になることを意識してし加水している。
(プランター受け皿に水が少し染み出るほどにして、溢れた水は翌日にプランターに戻す)
このように土壌水分はほぼ一定に保ってはいるはずだが、土壌表面放射線量の低下/上昇の変化のようすから、
水分量の過多だけによる変化ではないと言える。
② 土壌表面の測定器の位置が土壌面から離れることによって線量が弱まるものである。
確かに少しづつは離れていき、それによって線量が低下することは確かである。
しかし土壌表面の変化のようすをよく見ると、実験そのⅠでは大きく上昇している箇所もある。
実験そのⅡでは、ボカシなどの投入量に必ずしも比例しない放射線量の変化も随所に見られることから、
距離以外の要因もあるものと考える。
③ 土壌中のセシウムが徐々に底面方向に移動しているのではないか?
それならば、底面放射線が徐々に上昇して現象には合わないし、自然乾燥時の変化を見る限り、
セシウムの沈下は有意には表れてはいない。
以上のことから、土壌表面の表面線量の低下や変化は、土壌微生物の働きが深く関与しているものと考えられる。
なお、多様な微生物群のうち、光子エネルギーを吸収利用できる微生物は「光合成細菌」しかないことから、
放射線の土壌表面での低減は光合成細菌によるものに違いない考える(放射線吸収説)。
(2) 光合成細菌は多量に増殖している (放射線吸収説の裏付け-2)
 この写真は、実験そのⅡのプランター内壁面のものだが、
この写真は、実験そのⅡのプランター内壁面のものだが、光合成細菌(赤色)がびっしりと増殖し付着している。
壁面内側には太陽光の直接の照射はないので、
壁面の熱によって増殖がしたものと思われが、
底面にはそれもなく光合成細菌は全く増殖していない。
このような状況から、光合成細菌はプランター内でも増殖していることが確認できる。
土壌表面の光合成細菌は迂闊にも確認しなかったが、太陽光や反射光が最も多く直接照射する所なので、
内部壁面よりも遥かに多量の光合成細菌は層を形成していると考えられる。
(3) 放射線吸収説を裏付ける土壌撹拌後の放射線量の変化 (放射線吸収説の裏付け-3)
「実験そのⅡ」において、実験終盤で「プランターの土壌全体を撹拌した直後に測定」したところ、
放射線量が大巾に変化しているが、これは次のように説明できる。
① 土壌を撹拌したことで、表層に集中していた光合成細菌が土壌全体に散らばったので、
底面方向に移動した光合成細菌によって放射線は吸収され、結果として放射線量が低く測定された(E)
② 同じ理由により、表面に集中していた光合成細菌が大幅に減ったので、土壌放射線の吸収量が激減し、
表面の放射線量は大幅に上昇した(F)
この二つの現象は、光合成細菌による「放射線吸収説」を強く裏付けるものであり、物的証拠となるデータでもある。
(4) 二酸化炭素を供給すると光合成細菌が活性化する (放射線吸収説の裏付け-4)
実験そのⅠおよび実験そのⅡの土壌表面に変化を再度注視していただきたい。
酵母菌が増えるような資材を投入すると、土壌表面放射線が減少する、という因果関係な現象が読み取れる。
(実験では、土壌表面の放射線量を低下させるために意識的に酵母菌の活動を強化した)
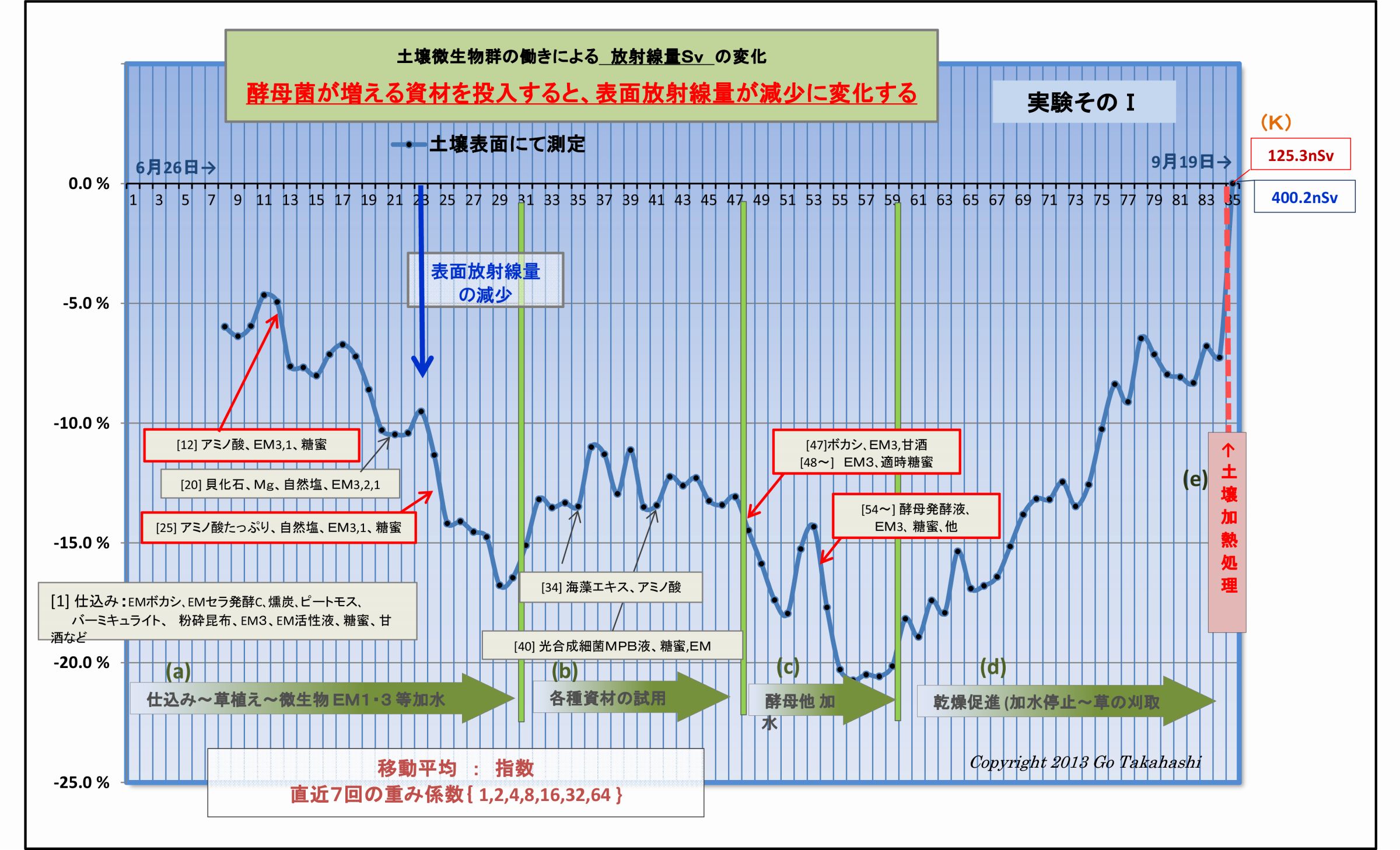
米ぬか発酵ボカシや麹に酵母菌が多く含まれており、糖蜜や甘酒(麹由来の)は酵母菌が増殖するための
優良なエサであり、甘さの残る酵母発酵液は酵母菌が多量に増殖している。
光合成細菌は酵母菌と相性がいい、酵母菌から発生する二酸化炭素は光合成細菌には「不可欠なエサ」であるからだ。
下図は、光合成細菌の生体内部の機能をまとめたものである。
光合成細菌は、二酸化炭素CO2と光子エネルギーをエサとして糖を産生する微生物である。
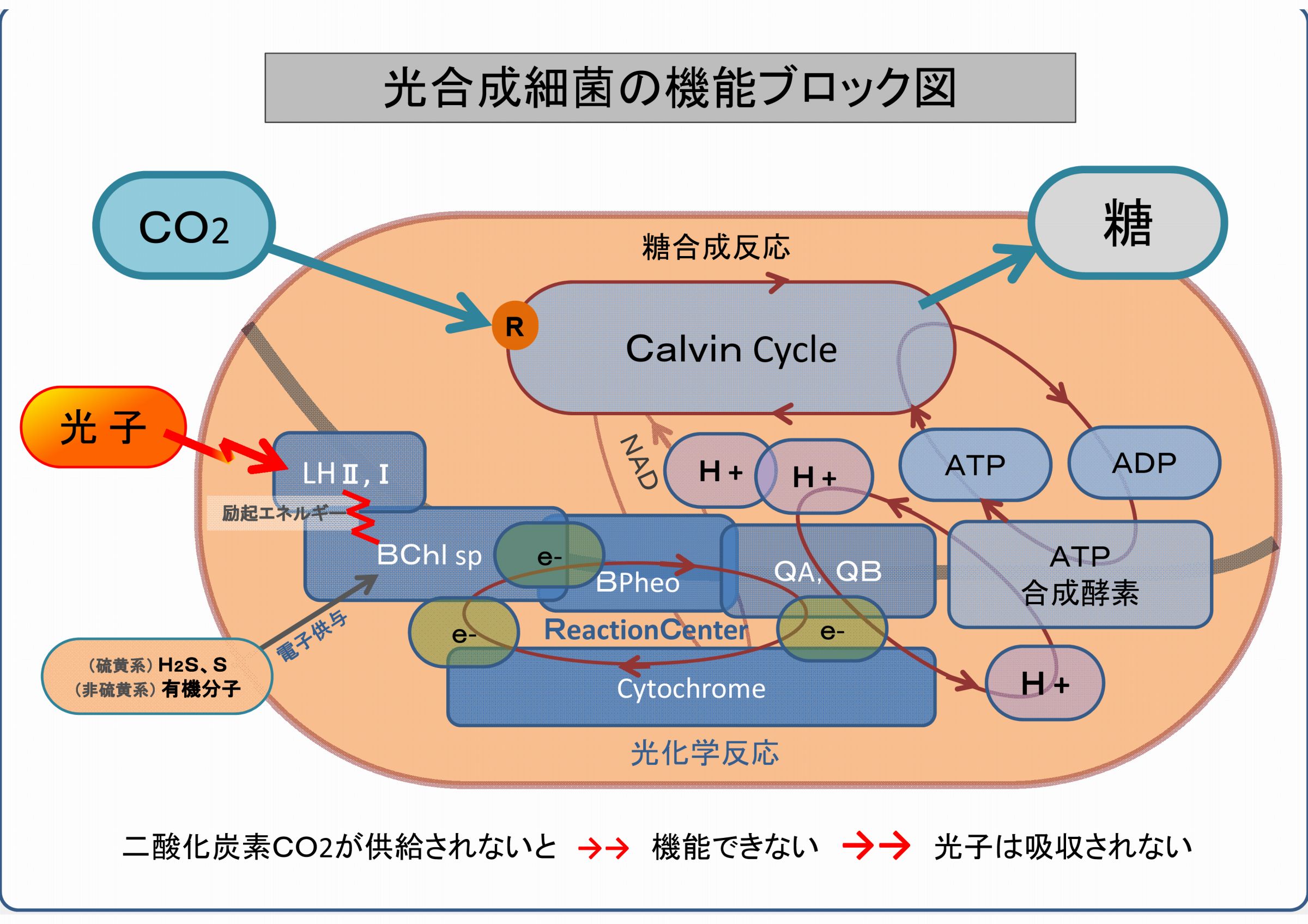
光合成細菌は、二酸化炭素CO2がなければ働けない・機能できない。
実験データによる土壌表面の放射線量の変化を注視すると、酵母関連資材を投入すると表面放射線量は低下し、
酵母菌が少ないと表面放射線は反転上昇するようすが見て取れる。
つまり、光合成細菌は、二酸化炭素CO2が供給されないと光子エネルギーを吸収できないのである。
このことを、ブロック図の機能を出口方向から順に説明する。
・CO2が無いと、糖を合成できないだけでなく、カルビン回路そのものが循環しない
↓↓↓↓
・ADPが還流されないので、ATP合成酵素は働けない
↓↓↓↓
・プロトンH+の循環が止まる(飽和状態)
↓↓↓↓
・e-電子の循環が止まる
↓↓↓↓
・シトクロームからの電子の還流が無いと酸化したバクテリオクロロフィルの機能は復元できない
↓↓↓↓
・光子からのエネルギーは吸収されない => 放射線は吸収できない
つまり、二酸化炭素の供給がなければ光合成細菌は光子(放射線)エネルギーを吸収できない、ということである。
( 身近な例だが、電力は柱上トランスの電磁誘導を経由して二次側の家庭に引き込まれているが、
電気が消費されなければ電流はながれないので、高圧線の電力は消費=吸収されない。
光合成細菌でいうなら、糖合成に必要なの原料が消費されないと光子エネルギーは吸収されない原理と同じである )
これが光合成細菌による「放射線吸収説」の正しさを説明できるメカニズムであり、
実験で投入した酵母促進資材の「不足/充足」と表面線量の「上昇/低下」の因果関係の説明にもなっている。
光合成細菌単独では働けない、光合成細菌を中核とした良好なマイクロバイオーム(微生物叢)が放射線の吸収・利用を高めている。